

Structure-function characterization of the human mitochondrial thiamin pyrophosphate transporter (hMTPPT; SLC25A19): Important roles for Ile(33), Ser(34), Asp(37), His(137) and Lys(291)
- PMID: 27188525
- PMCID: PMC4900926
- DOI: 10.1016/j.bbamem.2016.05.011
Structure-function characterization of the human mitochondrial thiamin pyrophosphate transporter (hMTPPT; SLC25A19): Important roles for Ile(33), Ser(34), Asp(37), His(137) and Lys(291)
Abstract
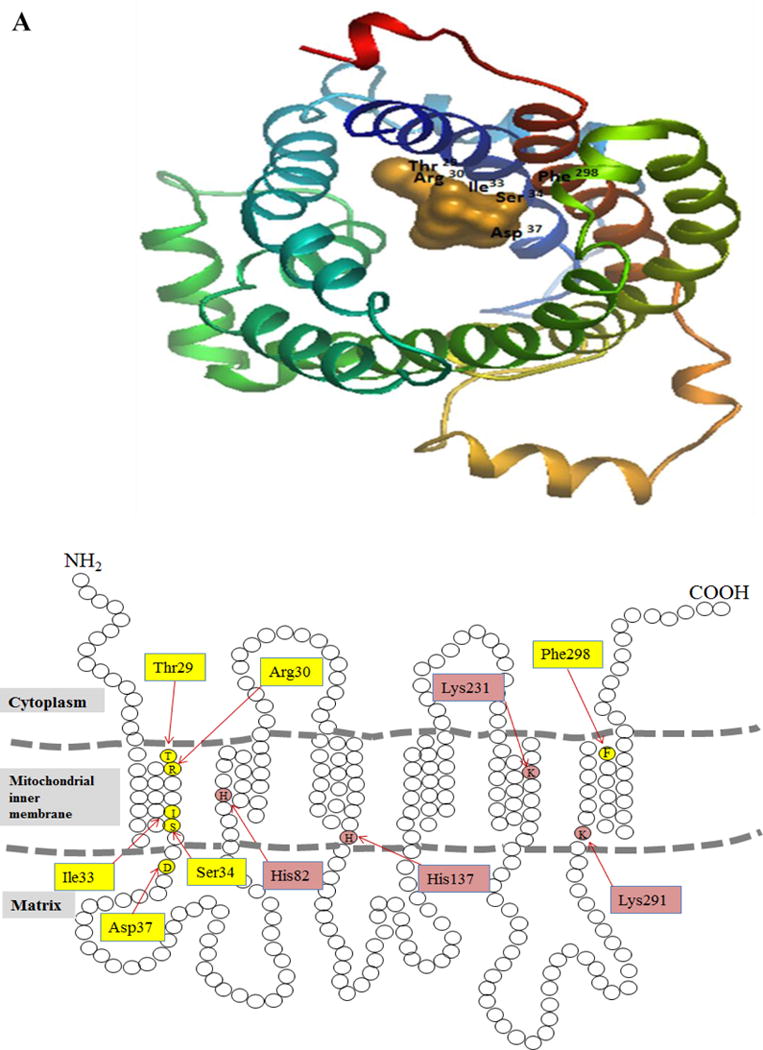
Thiamin plays a critical role in cellular energy metabolism. Mammalian cells obtain the vitamin from their surroundings, converted it to thiamin pyrophosphate (TPP) in the cytoplasm, followed by uptake of TPP by mitochondria via a carrier-mediated process that involves the MTPPT (product of the SLC25A19 gene). Previous studies have characterized different physiological/biological aspects of the human MTPPT (hMTPPT), but less is known about structural features that are important for its function. Here, we used a protein-docking model ("Phyre2" and "DockingServer") to predict residues that may be important for function (substrate recognition) of the hMTPPT; we also examined the role of conserved positively-charged residues predicted ("PRALINE") to be in the trans-membrane domains (TMDs) in uptake of the negatively-charged TPP. Among the six residues predicted by the docking model (i.e., Thr(29), Arg(30), Ile(33), Ser(34), Asp(37) and Phe(298)), only Ile(33), Ser(34) and Asp(37) were found to be critical for function. While no change in translational efficiency/protein stability of the Ser(34) mutant was observed, both the Ile(33) and Asp(37) mutants showed a decrease in this parameter(s); there was also a decrease in the expression of the latter two mutants in mitochondria. A need for a polar residue at position 34 of the hMTPPT was evident. Our findings with the positively-charged residues (i.e., His(82), His(137), Lys(231) and Lys(291)) predicted in the TMD showed that His(137) and Lys(291) are important for function (via a role in proper delivery of the protein to mitochondria). These investigations provide important information about the structure-function relationship of the hMTPPT.
Keywords: Mitochondria; Multiple sequence alignment; Protein-docking modeling; Site-directed mutagenesis; Thiamin pyrophosphate transporter; Uptake.
Copyright © 2016 Elsevier B.V. All rights reserved.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures




References
-
- Berdanier CD. Advanced Nutrition-Micronutrients. CRC Press; New York: 1998. pp. 80–88.
-
- Singleton CK, Martin PR. Molecular mechanisms of thiamin utilization. Curr Mol Med. 2001;1:197–207. - PubMed
-
- Calingasan N, Chun W, Park L, Uchida K, Gibson GE. Oxidative stress is associated with region-specific neuronal death during thiamine deficiency. J Neuropathol Exp Neurol. 1999;58:946–958. - PubMed
-
- Portari GV, Marchini JS, Vannucchi H, Jordao AA. Antioxidant effect of thiamine on acutely alcoholized rats and lack of efficacy using thiamine or glucose to reduce blood alcohol content. Basic Clin Pharmacol Toxicol. 2008;103:482–486. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
