

Identification of Diverse Mycoviruses through Metatranscriptomics Characterization of the Viromes of Five Major Fungal Plant Pathogens
- PMID: 27194764
- PMCID: PMC4944287
- DOI: 10.1128/JVI.00357-16
Identification of Diverse Mycoviruses through Metatranscriptomics Characterization of the Viromes of Five Major Fungal Plant Pathogens
Abstract
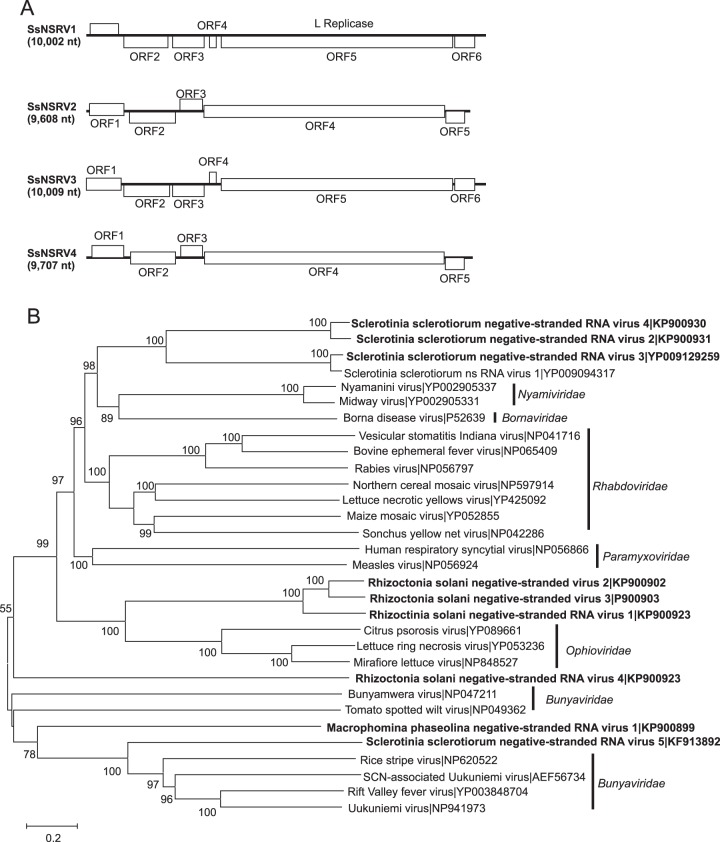
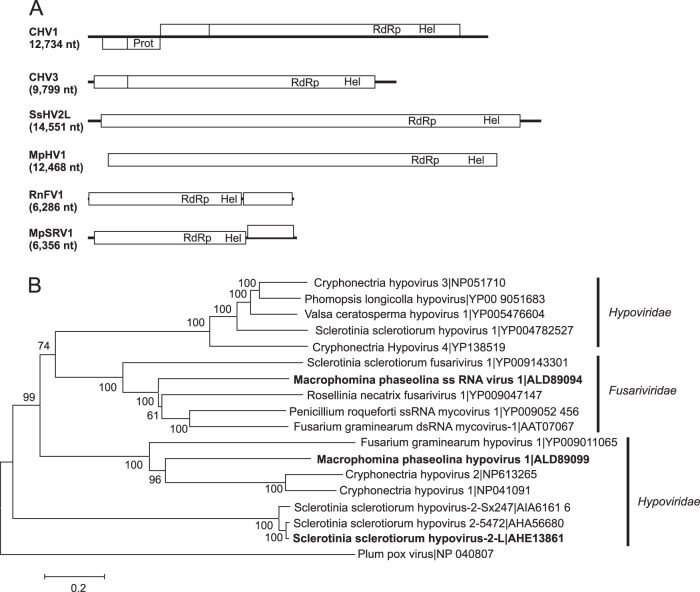
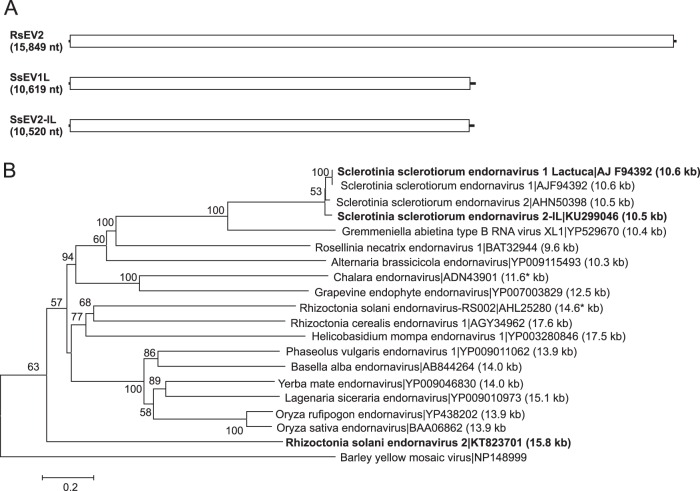
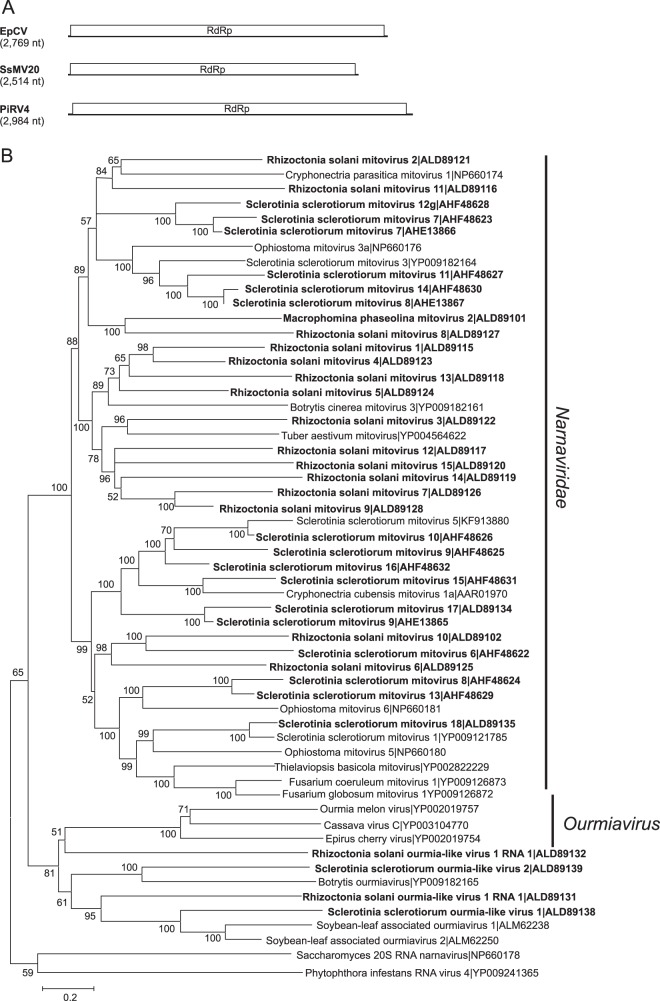
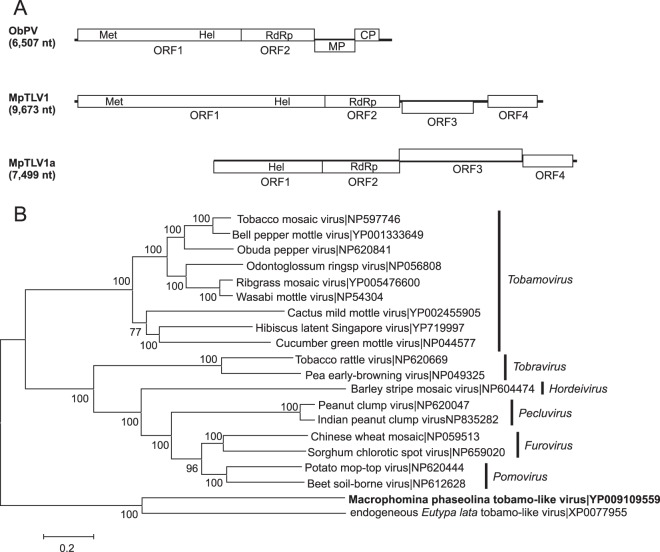
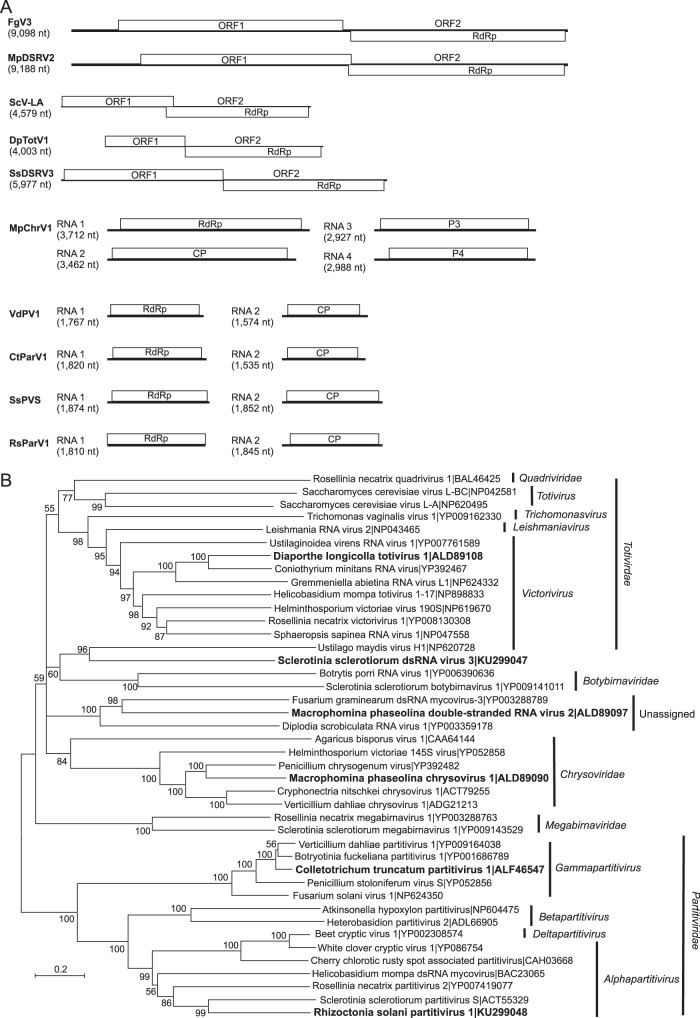
Mycoviruses can have a marked effect on natural fungal communities and influence plant health and productivity. However, a comprehensive picture of mycoviral diversity is still lacking. To characterize the viromes of five widely dispersed plant-pathogenic fungi, Colletotrichum truncatum, Macrophomina phaseolina, Diaporthe longicolla, Rhizoctonia solani, and Sclerotinia sclerotiorum, a high-throughput sequencing-based metatranscriptomic approach was used to detect viral sequences. Total RNA and double-stranded RNA (dsRNA) from mycelia and RNA from samples enriched for virus particles were sequenced. Sequence data were assembled de novo, and contigs with predicted amino acid sequence similarities to viruses in the nonredundant protein database were selected. The analysis identified 72 partial or complete genome segments representing 66 previously undescribed mycoviruses. Using primers specific for each viral contig, at least one fungal isolate was identified that contained each virus. The novel mycoviruses showed affinity with 15 distinct lineages: Barnaviridae, Benyviridae, Chrysoviridae, Endornaviridae, Fusariviridae, Hypoviridae, Mononegavirales, Narnaviridae, Ophioviridae, Ourmiavirus, Partitiviridae, Tombusviridae, Totiviridae, Tymoviridae, and Virgaviridae More than half of the viral sequences were predicted to be members of the Mitovirus genus in the family Narnaviridae, which replicate within mitochondria. Five viral sequences showed strong affinity with three families (Benyviridae, Ophioviridae, and Virgaviridae) that previously contained no mycovirus species. The genomic information provides insight into the diversity and taxonomy of mycoviruses and coevolution of mycoviruses and their fungal hosts.
Importance: Plant-pathogenic fungi reduce crop yields, which affects food security worldwide. Plant host resistance is considered a sustainable disease management option but may often be incomplete or lacking for some crops to certain fungal pathogens or strains. In addition, the rising issues of fungicide resistance demand alternative strategies to reduce the negative impacts of fungal pathogens. Those fungus-infecting viruses (mycoviruses) that attenuate fungal virulence may be welcome additions for mitigation of plant diseases. By high-throughput sequencing of the RNAs from 275 isolates of five fungal plant pathogens, 66 previously undescribed mycoviruses were identified. In addition to identifying new potential biological control agents, these results expand the grand view of the diversity of mycoviruses and provide possible insights into the importance of intracellular and extracellular transmission in fungus-virus coevolution.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures


Similar articles
-
Survey of Early-Diverging Lineages of Fungi Reveals Abundant and Diverse Mycoviruses.mBio. 2020 Sep 8;11(5):e02027-20. doi: 10.1128/mBio.02027-20. mBio. 2020. PMID: 32900807 Free PMC article.
-
Deep Sequencing Analysis Reveals the Mycoviral Diversity of the Virome of an Avirulent Isolate of Rhizoctonia solani AG-2-2 IV.PLoS One. 2016 Nov 4;11(11):e0165965. doi: 10.1371/journal.pone.0165965. eCollection 2016. PLoS One. 2016. PMID: 27814394 Free PMC article.
-
Divergent RNA viruses in Macrophomina phaseolina exhibit potential as virocontrol agents.Virus Evol. 2020 Dec 18;7(1):veaa095. doi: 10.1093/ve/veaa095. eCollection 2021 Jan. Virus Evol. 2020. PMID: 33505706 Free PMC article.
-
Viruses Infecting the Plant Pathogenic Fungus Rhizoctonia solani.Viruses. 2019 Nov 30;11(12):1113. doi: 10.3390/v11121113. Viruses. 2019. PMID: 31801308 Free PMC article. Review.
-
[Mycoviruses and importance in mycology].Mikrobiyol Bul. 2017 Oct;51(4):404-412. doi: 10.5578/mb.54128. Mikrobiyol Bul. 2017. PMID: 29153071 Review. Turkish.
Cited by
-
Armillaria root rot fungi host single-stranded RNA viruses.Sci Rep. 2021 Apr 1;11(1):7336. doi: 10.1038/s41598-021-86343-7. Sci Rep. 2021. PMID: 33795735 Free PMC article.
-
Greetings from virologists to mycologists: A review outlining viruses that live in fungi.Mycoscience. 2024 Jan 31;65(1):1-11. doi: 10.47371/mycosci.2023.11.004. eCollection 2024. Mycoscience. 2024. PMID: 39239117 Free PMC article.
-
The Expanding Mycovirome of Aspergilli.J Fungi (Basel). 2024 Aug 17;10(8):585. doi: 10.3390/jof10080585. J Fungi (Basel). 2024. PMID: 39194910 Free PMC article. Review.
-
Extreme Diversity of Mycoviruses Present in Isolates of Rhizoctonia solani AG2-2 LP From Zoysia japonica From Brazil.Front Cell Infect Microbiol. 2019 Jul 12;9:244. doi: 10.3389/fcimb.2019.00244. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31355150 Free PMC article.
-
Characterization of the Mycovirome from the Plant-Pathogenic Fungus Cercospora beticola.Viruses. 2021 Sep 24;13(10):1915. doi: 10.3390/v13101915. Viruses. 2021. PMID: 34696345 Free PMC article.
References
-
- Wu ZQ, Ren XW, Yang L, Hu YF, Yang J, He GM, Zhang JP, Dong J, Sun LL, Du J, Liu LG, Xue Y, Wang JM, Yang F, Zhang SY, Jin Q. 2012. Virome analysis for identification of novel mammalian viruses in bat species from Chinese provinces. J Virol 86:10999–11012. doi: 10.1128/JVI.01394-12. - DOI - PMC - PubMed
-
- Cook S, Chung BYW, Bass D, Moureau G, Tang SY, McAlister E, Culverwell CL, Glucksman E, Wang H, Brown TDK, Gould EA, Harbach RE, de Lamballerie X, Firth AE. 2013. Novel virus discovery and genome reconstruction from field RNA samples reveals highly divergent viruses in Dipteran hosts. PLoS One 8:e80720. doi: 10.1371/journal.pone.0080720. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
