

Venus flytrap carnivorous lifestyle builds on herbivore defense strategies
- PMID: 27197216
- PMCID: PMC4889972
- DOI: 10.1101/gr.202200.115
Venus flytrap carnivorous lifestyle builds on herbivore defense strategies
Abstract
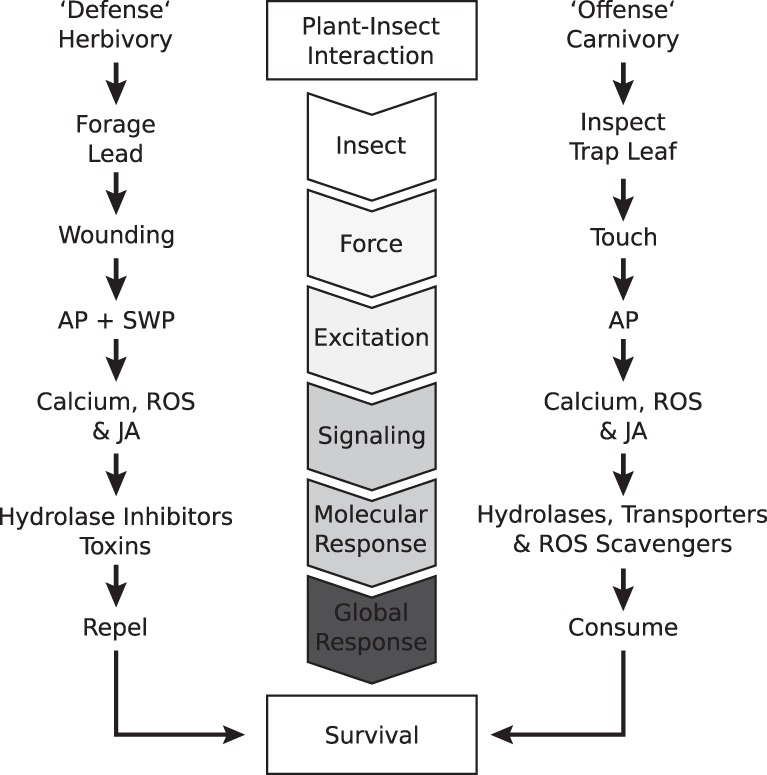
Although the concept of botanical carnivory has been known since Darwin's time, the molecular mechanisms that allow animal feeding remain unknown, primarily due to a complete lack of genomic information. Here, we show that the transcriptomic landscape of the Dionaea trap is dramatically shifted toward signal transduction and nutrient transport upon insect feeding, with touch hormone signaling and protein secretion prevailing. At the same time, a massive induction of general defense responses is accompanied by the repression of cell death-related genes/processes. We hypothesize that the carnivory syndrome of Dionaea evolved by exaptation of ancient defense pathways, replacing cell death with nutrient acquisition.
© 2016 Bemm et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Adamec L. 1997. Mineral nutrition of carnivorous plants: a review. Bot Rev 63: 273–299.
-
- Adlassnig W, Koller-Peroutka M, Bauer S, Koshkin E, Lendl T, Lichtscheidl IK. 2012. Endocytotic uptake of nutrients in carnivorous plants. Plant J 71: 303–313. - PubMed
-
- Albert V, Williams S, Chase M. 1992. Carnivorous plants: phylogeny and structural evolution. Science 257: 1491–1495. - PubMed
-
- Alexa A, Rahnenfuhrer J, Lengauer T. 2006. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 22: 1600–1607. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources