

A hot L1 retrotransposon evades somatic repression and initiates human colorectal cancer
- PMID: 27197217
- PMCID: PMC4889970
- DOI: 10.1101/gr.201814.115
A hot L1 retrotransposon evades somatic repression and initiates human colorectal cancer
Abstract
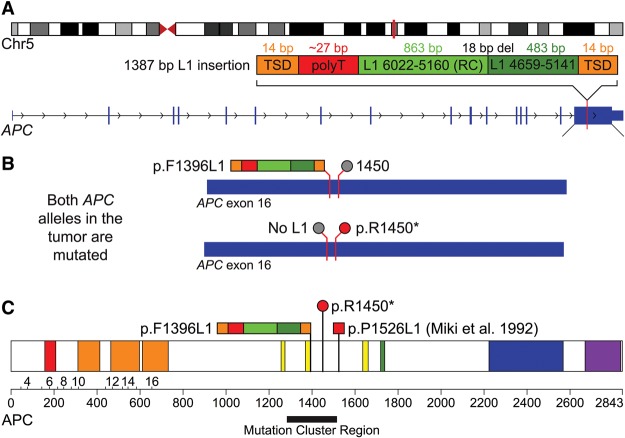
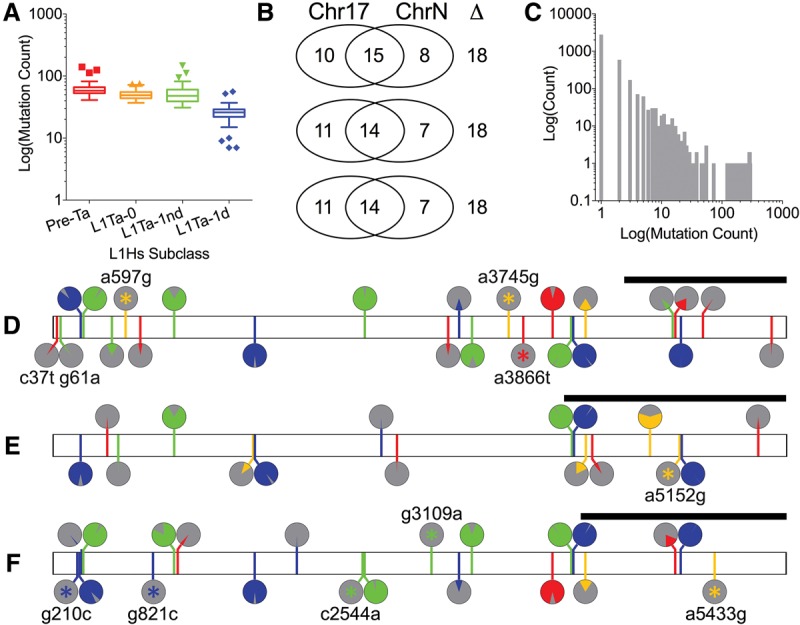
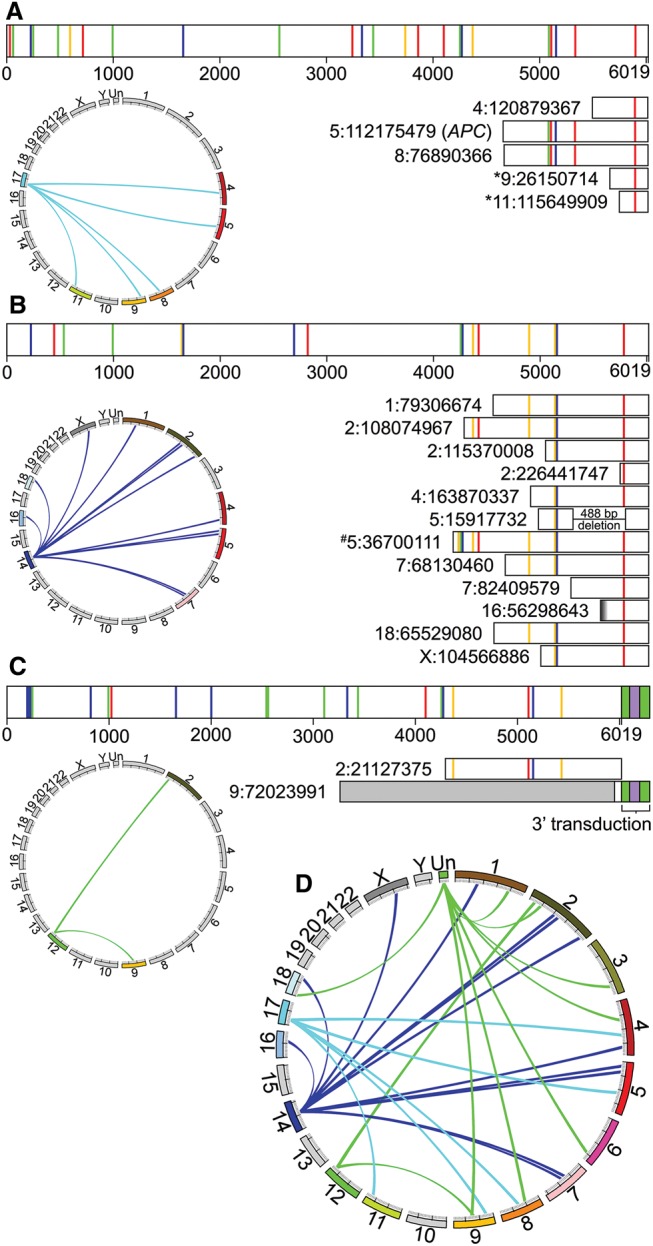
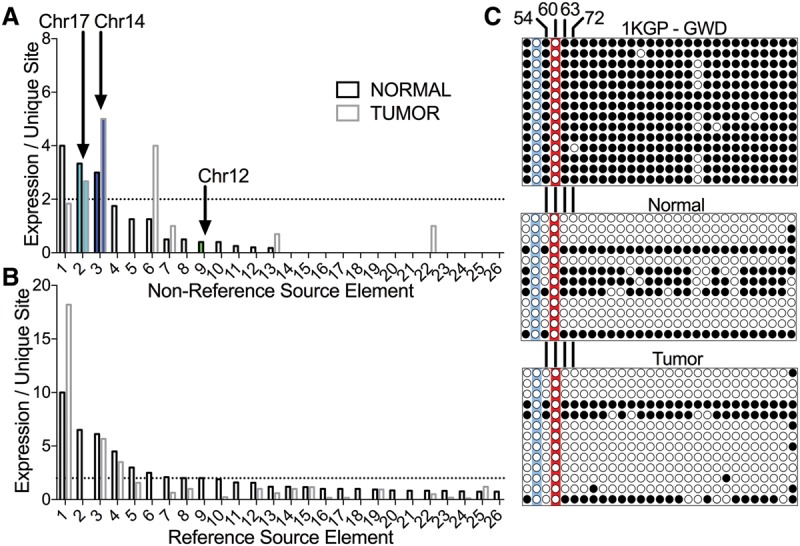
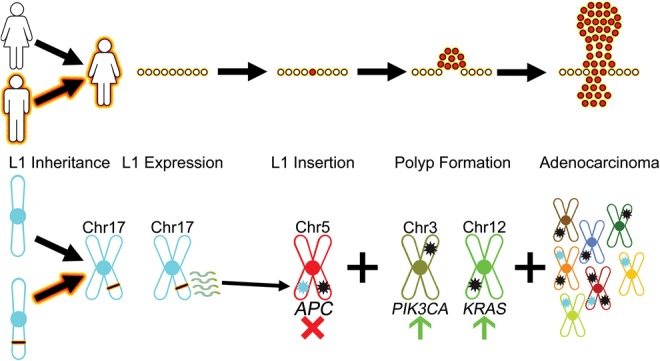
Although human LINE-1 (L1) elements are actively mobilized in many cancers, a role for somatic L1 retrotransposition in tumor initiation has not been conclusively demonstrated. Here, we identify a novel somatic L1 insertion in the APC tumor suppressor gene that provided us with a unique opportunity to determine whether such insertions can actually initiate colorectal cancer (CRC), and if so, how this might occur. Our data support a model whereby a hot L1 source element on Chromosome 17 of the patient's genome evaded somatic repression in normal colon tissues and thereby initiated CRC by mutating the APC gene. This insertion worked together with a point mutation in the second APC allele to initiate tumorigenesis through the classic two-hit CRC pathway. We also show that L1 source profiles vary considerably depending on the ancestry of an individual, and that population-specific hot L1 elements represent a novel form of cancer risk.
© 2016 Scott et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Boissinot S, Chevret P, Furano AV. 2000. L1 (LINE-1) retrotransposon evolution and amplification in recent human history. Mol Biol Evol 17: 915–928. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical