

doi: 10.1186/s12915-016-0261-6.
What is memory? The present state of the engram
Affiliations
- PMID: 27197636
- PMCID: PMC4874022
- DOI: 10.1186/s12915-016-0261-6
Item in Clipboard
What is memory? The present state of the engram
BMC Biol.
.
Abstract
The mechanism of memory remains one of the great unsolved problems of biology. Grappling with the question more than a hundred years ago, the German zoologist Richard Semon formulated the concept of the engram, lasting connections in the brain that result from simultaneous "excitations", whose precise physical nature and consequences were out of reach of the biology of his day. Neuroscientists now have the knowledge and tools to tackle this question, however, and this Forum brings together leading contemporary views on the mechanisms of memory and what the engram means today.
Figures
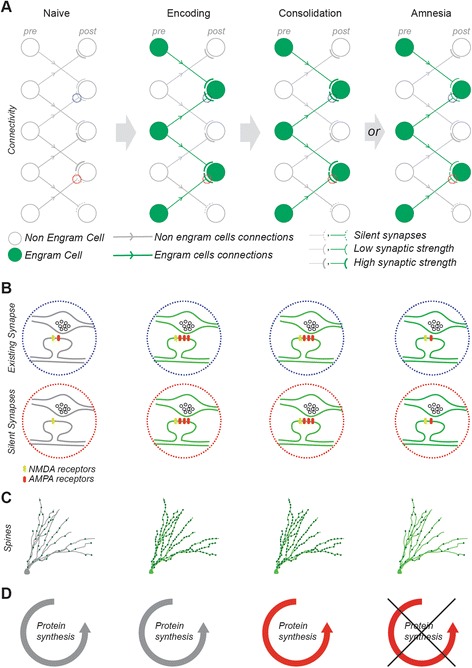
Synaptic connectivity between engram cells as a mechanism for memory storage. a Cellular connectivity in a feedforward excitatory circuit, b synaptic configuration, c dendritic spine density, and d protein synthesis state, shown in a naïve circuit, a circuit during encoding, a circuit after consolidation, or a circuit in an amnesic condition. Engram circuit, cells, and synapses are displayed in green, non-engram in gray. In the naïve state, the circuit displays a variety of synaptic patterns, including strong (thick gray lines) and weak synapses (thin gray lines) as well as silent synapses (dotted lines) exclusively expressing NMDA receptors. During encoding, a network of engram cells is recruited. The preferential connection between engram cells occurs either by potentiation of existing connections (blue dotted circles) or by unsilencing synapses (red dotted circles). A spine density increase supports the synaptic changes. During consolidation, the steady state synthesis of AMPA receptors is shifted to a higher level and the disruption of consolidation with protein synthesis inhibitors (PSI) results in retrograde amnesia. However, during PSI-induced amnesia, memory storage persists within an engram-specific set of weak synaptic connections
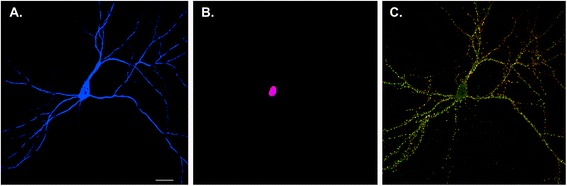
Neuron versus synaptic maps of memory. The Golgi method used by Ramon y Cajal at the turn of the 19th century revealed that the brain consisted of individual nerve cells, leading to the formulation of the neuron doctrine. In modern neurobiology, expression of soluble fluorescent proteins allows visualization of individual neurons in the brain or, as shown here, in a hippocampal pyramidal neuron in dissociated culture (blue) (a). Identification of immediate early genes, such as cFos, provides a means of labeling the nuclei of individual neurons that are activated to undergo transcription following neuronal activity (violet) (b). This provides a neuron-level map of activity. However, each neuron forms thousands of synapses, shown in c by labeling a single neuron with presynaptic (green) and postsynaptic (red) markers (yellow where the pre- and post-synaptic elements are adjacent to one another). Note that this neuron is connected to other neurons in the culture that are not shown. Developing a synaptic rather than a neuron-wide map of memory requires obtaining activity-dependent markers that label the activated synapses rather than nuclei. Understanding synaptic maps will not only require understanding the identity of individual neurons but also the details of their cell-to-cell interactions
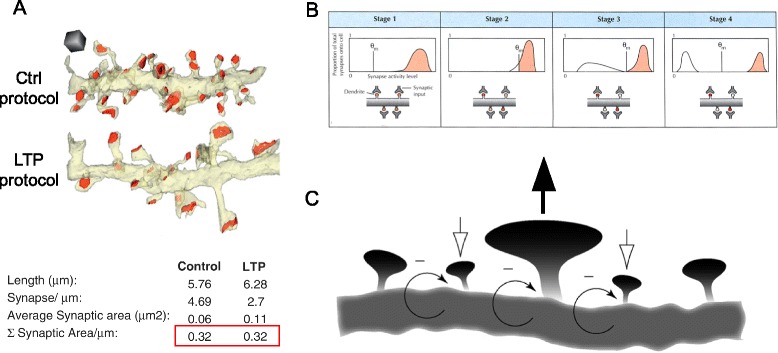
Possible mechanisms of synaptic modification in memory storage. a Structural synaptic scaling is analyzed by serial section electron microscopy in hippocampal tissue under control and LTP (induced via theta burst stimulation) conditions. Control and LTP dendrites have equal postsynaptic areas (red outline) despite differences in synaptic density and size. Adapted from [72], with permission. b Representation of metaplasticity occurring, shifting the plasticity threshold (θ
m). The graphs on the top row depict the distribution of synaptic responses, red shading indicating LTP and white shading LTD. The four synapses illustrated (bottom row) reflect how the shifting of θm would impact the strength of individual synapses (intensity of red shading correlating to strength). Stage 1 depicts a saturating event, where all of the synaptic strengths have been excited to levels above θm. Stage 2 shows a metaplastic response to the activity levels reached in stage 1. In stage 2, θm has shifted such that some synapses are below the new threshold and weaken in stage 3. This changed θm leads to a new stable state, illustrated in stage 4. Reproduced from [75], with permission. c Synaptic potentiation at one spine (center, upward-pointing black arrow added for emphasis) is predicted to temporarily increase activity at adjacent spines. In response to the increased activity, homeostatic plasticity weakens the entire area (negative feedback arrows), with closely neighboring spines being disproportionately weakened (downward-pointing white arrows). The weakened spines allow the overall dendritic length to maintain a constant level of activity with the central spine still maintaining a level of potentiation. Adapted from [76], with permission
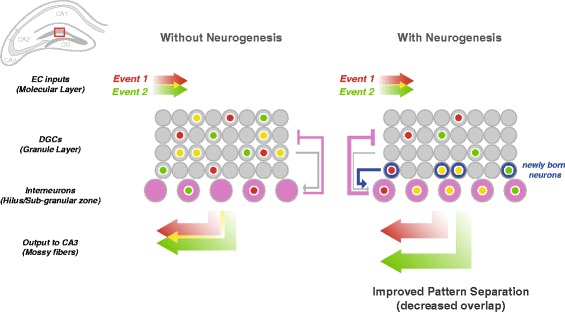
Immature adult-born neurons improve pattern-separation in the DG by enhancing feedback inhibition. Two events are encoded in separate but partially overlapping populations of activated DGCs in the DG (red and green, with overlap in yellow). DGCs receive strong inhibitory inputs from interneurons (purple) in the hilus and sub-granular zone. It is hypothesized that hyperactive immature adult-born DGCs (blue) drive these interneurons, enhancing feedback inhibition from the hilus, which results in decreased overlap of activated DGCs and output to CA3, thereby improving pattern separation
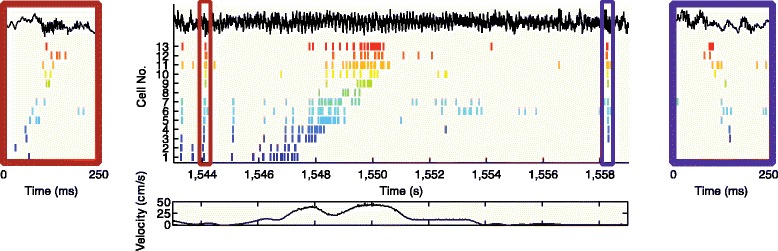
Place cell sequences experienced during behavior (middle panel) are replayed in both forward (left panel) and reverse (right panel) direction during awake SPW-R. The rat is moving from left to right on a familiar track. Spike trains for place fields of 13 CA3 pyramidal cells (color ticks, spikes of individual neurons) on the track are shown before (forward replay; left red box), during (middle), and after (reverse replay; right blue box) a single traversal. The CA1 local field potential is shown on top (black traces) and the animal’s velocity is shown below. Reproduced from [137]
References
-
- Hebb DO. The organization of behavior; a neuropsychological theory. New York: Wiley; 1949.
-
- Semon RW. The mneme. 1921.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
