

Orthology Analysis and In Vivo Complementation Studies to Elucidate the Role of DIR1 during Systemic Acquired Resistance in Arabidopsis thaliana and Cucumis sativus
- PMID: 27200039
- PMCID: PMC4854023
- DOI: 10.3389/fpls.2016.00566
Orthology Analysis and In Vivo Complementation Studies to Elucidate the Role of DIR1 during Systemic Acquired Resistance in Arabidopsis thaliana and Cucumis sativus
Erratum in
-
Corrigendum: Orthology Analysis and In Vivo Complementation Studies to Elucidate the Role of DIR1 during Systemic Acquired Resistance in Arabidopsis thaliana and Cucumis sativus.Front Plant Sci. 2018 Apr 12;9:460. doi: 10.3389/fpls.2018.00460. eCollection 2018. Front Plant Sci. 2018. PMID: 29681918 Free PMC article.
Abstract
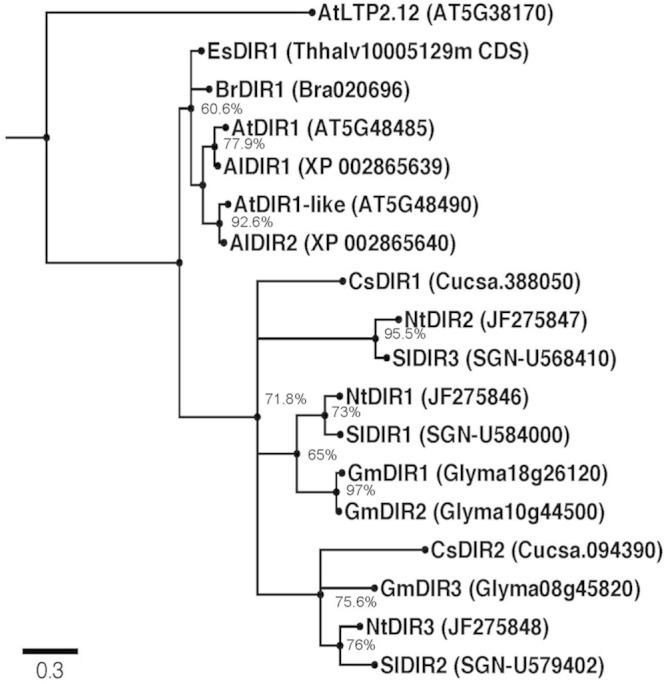
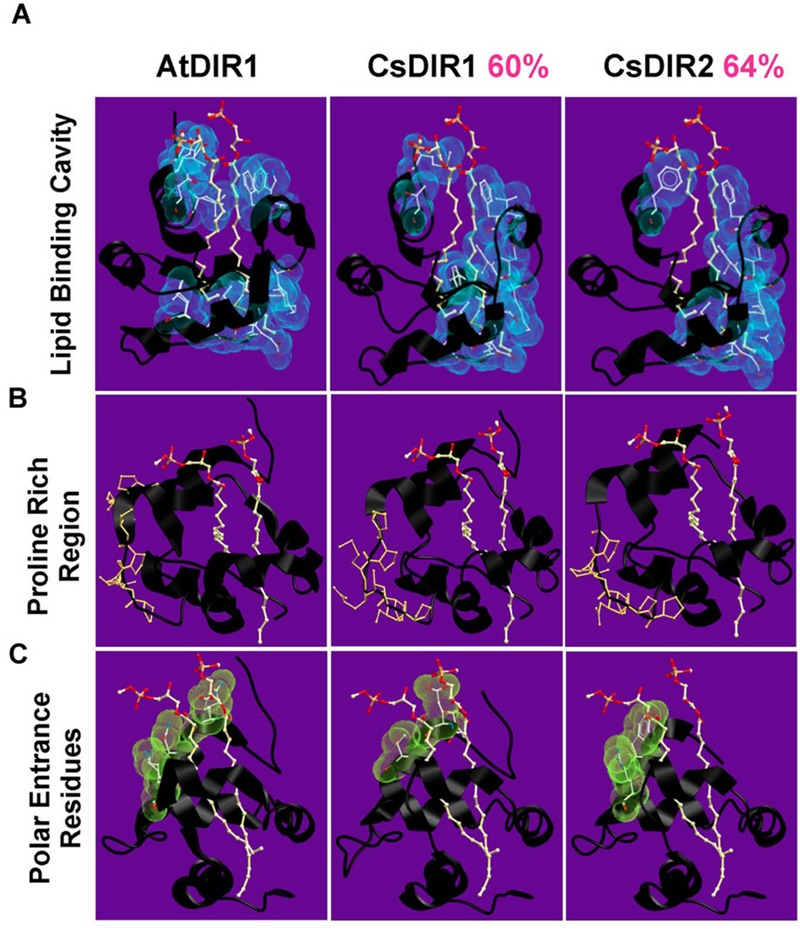
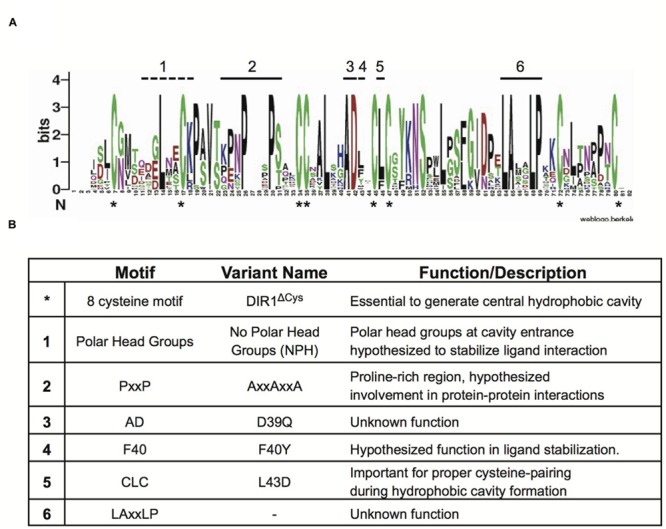
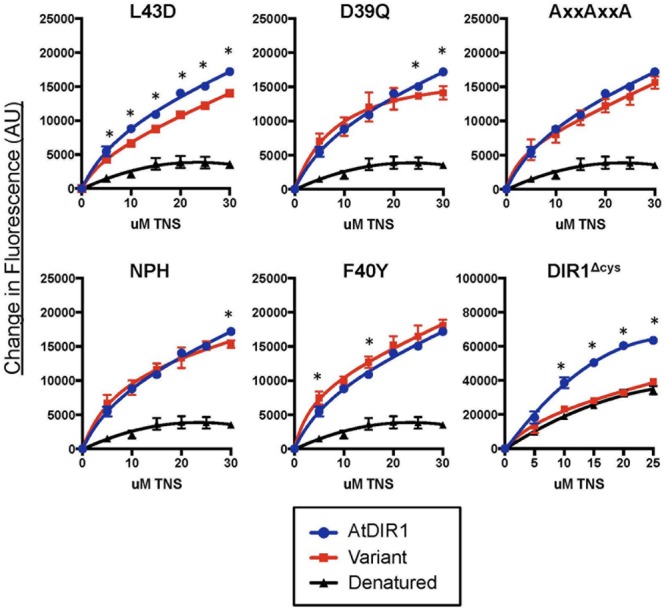
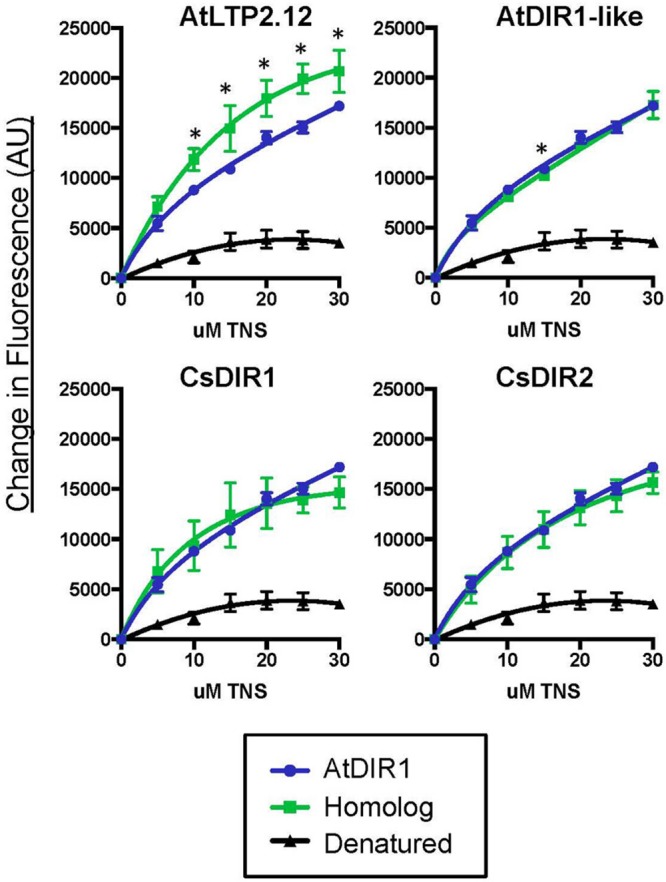
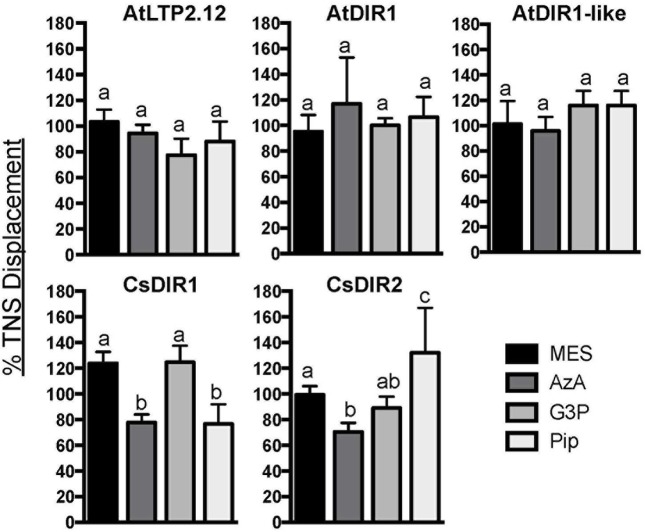
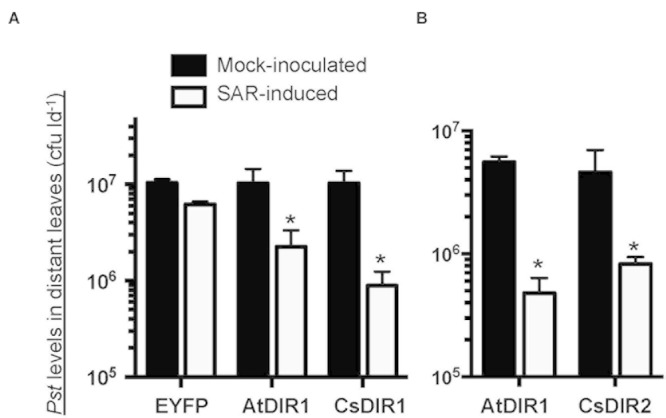
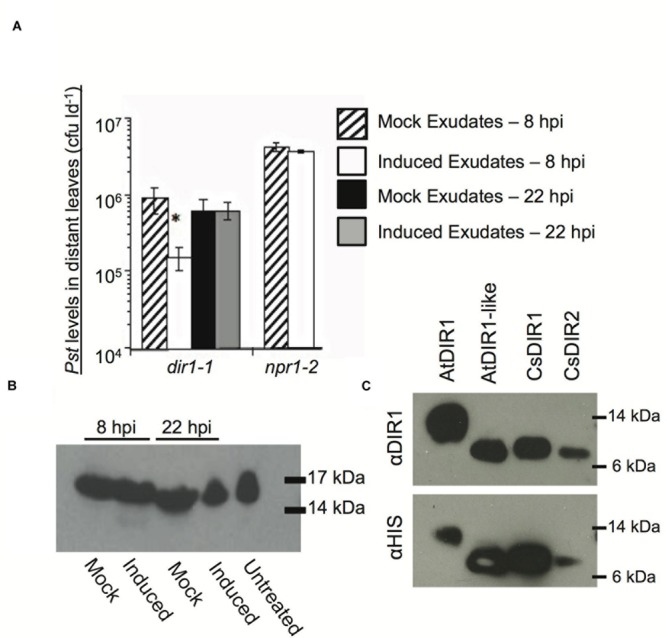
AtDIR1 (Defective in Induced Resistance1) is an acidic lipid transfer protein essential for systemic acquired resistance (SAR) in Arabidopsis thaliana. Upon SAR induction, DIR1 moves from locally infected to distant uninfected leaves to activate defense priming; however, a molecular function for DIR1 has not been elucidated. Bioinformatic analysis and in silico homology modeling identified putative AtDIR1 orthologs in crop species, revealing conserved protein motifs within and outside of DIR1's central hydrophobic cavity. In vitro assays to compare the capacity of recombinant AtDIR1 and targeted AtDIR1-variant proteins to bind the lipophilic probe TNS (6,P-toluidinylnaphthalene-2-sulfonate) provided evidence that conserved leucine 43 and aspartic acid 39 contribute to the size of the DIR1 hydrophobic cavity and possibly hydrophobic ligand binding. An Arabidopsis-cucumber SAR model was developed to investigate the conservation of DIR1 function in cucumber (Cucumis sativus), and we demonstrated that phloem exudates from SAR-induced cucumber rescued the SAR defect in the Arabidopsis dir1-1 mutant. Additionally, an AtDIR1 antibody detected a protein of the same size as AtDIR1 in SAR-induced cucumber phloem exudates, providing evidence that DIR1 function during SAR is conserved in Arabidopsis and cucumber. In vitro TNS displacement assays demonstrated that recombinant AtDIR1 did not bind the SAR signals azelaic acid (AzA), glycerol-3-phosphate or pipecolic acid. However, recombinant CsDIR1 and CsDIR2 interacted weakly with AzA and pipecolic acid. Bioinformatic and functional analyses using the Arabidopsis-cucumber SAR model provide evidence that DIR1 orthologs exist in tobacco, tomato, cucumber, and soybean, and that DIR1-mediated SAR signaling is conserved in Arabidopsis and cucumber.
Keywords: DIR1; cucumber; hydrophobic cavity; lipid transfer protein; long-distance signaling; systemic acquired resistance.
Figures
References
-
- Cameron R. K., Dixon R. A., Lamb C. J. (1994). Biologically induced systemic acquired resistance in Arabidopsis thaliana. Plant J. 5 715–725. 10.1111/j.1365-313X.1994.00715.x - DOI
-
- Cameron R. K., Paiva N. L., Lamb C. J., Dixon R. A. (1999). Accumulation of salicylic acid and PR-1 gene transcripts in relation to the systemic acquired resistance (SAR) response induced by Pseudomonas syringae pv. tomato in Arabidopsis. Phys. Mol. Plant Pathol. 55 121–130. 10.1006/pmpp.1999.0214 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous