

Integrin-Linked Kinase in Muscle Is Necessary for the Development of Insulin Resistance in Diet-Induced Obese Mice
- PMID: 27207548
- PMCID: PMC4878430
- DOI: 10.2337/db15-1434
Integrin-Linked Kinase in Muscle Is Necessary for the Development of Insulin Resistance in Diet-Induced Obese Mice
Abstract
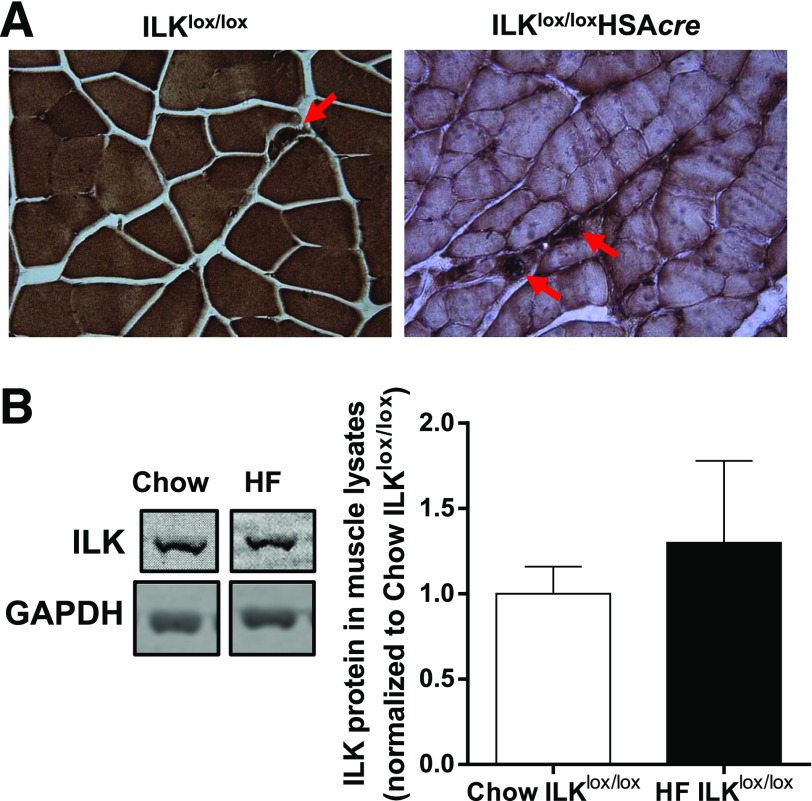
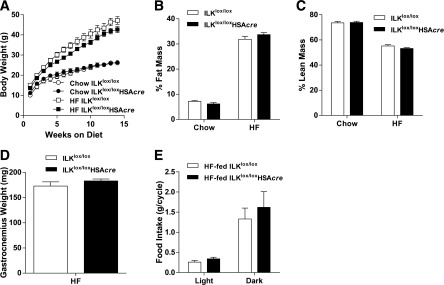
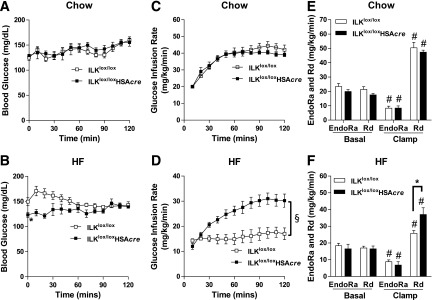
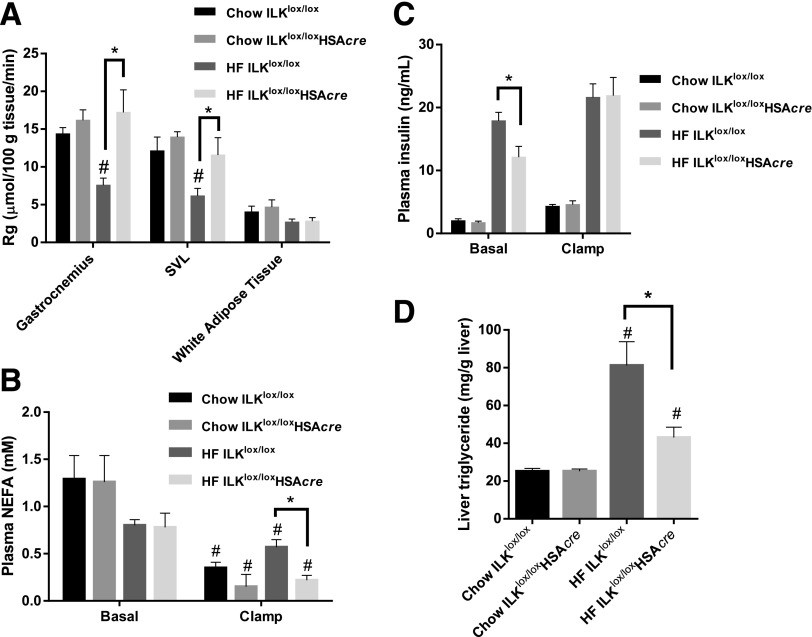
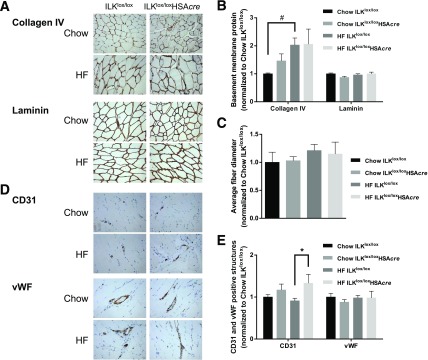
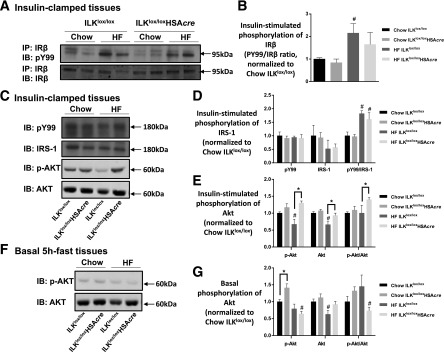
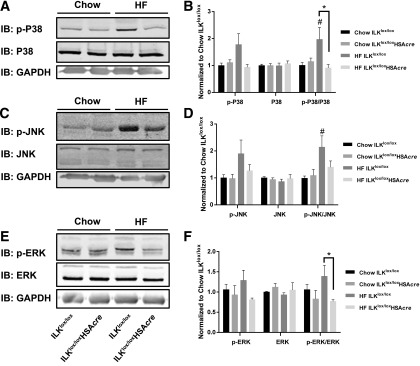
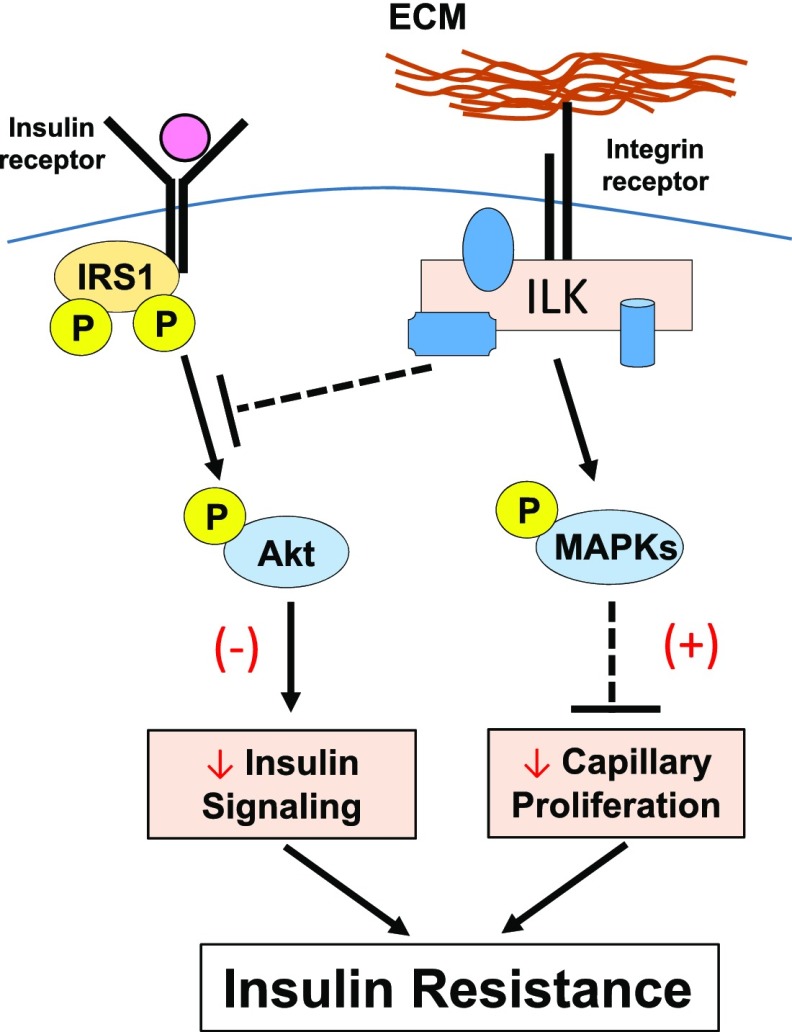
Diet-induced muscle insulin resistance is associated with expansion of extracellular matrix (ECM) components, such as collagens, and the expression of collagen-binding integrin, α2β1. Integrins transduce signals from ECM via their cytoplasmic domains, which bind to intracellular integrin-binding proteins. The integrin-linked kinase (ILK)-PINCH-parvin (IPP) complex interacts with the cytoplasmic domain of β-integrin subunits and is critical for integrin signaling. In this study we defined the role of ILK, a key component of the IPP complex, in diet-induced muscle insulin resistance. Wild-type (ILK(lox/lox)) and muscle-specific ILK-deficient (ILK(lox/lox)HSAcre) mice were fed chow or a high-fat (HF) diet for 16 weeks. Body weight was not different between ILK(lox/lox) and ILK(lox/lox)HSAcre mice. However, HF-fed ILK(lox/lox)HSAcre mice had improved muscle insulin sensitivity relative to HF-fed ILK(lox/lox) mice, as shown by increased rates of glucose infusion, glucose disappearance, and muscle glucose uptake during a hyperinsulinemic-euglycemic clamp. Improved muscle insulin action in the HF-fed ILK(lox/lox)HSAcre mice was associated with increased insulin-stimulated phosphorylation of Akt and increased muscle capillarization. These results suggest that ILK expression in muscle is a critical component of diet-induced insulin resistance, which possibly acts by impairing insulin signaling and insulin perfusion through capillaries.
© 2016 by the American Diabetes Association. Readers may use this article as long as the work is properly cited, the use is educational and not for profit, and the work is not altered.
Figures
References
-
- Reutrakul S, Van Cauter E. Interactions between sleep, circadian function, and glucose metabolism: implications for risk and severity of diabetes. Ann N Y Acad Sci 2014;1311:151–173 - PubMed
-
- Inzucchi SE, Bergenstal RM, Buse JB, et al. . Management of hyperglycaemia in type 2 diabetes: a patient-centered approach. Position statement of the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetologia 2012;55:1577–1596 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
