

Editor's Highlight: Hydroxyurea Exposure Activates the P53 Signaling Pathway in Murine Organogenesis-Stage Embryos
- PMID: 27208086
- PMCID: PMC4960909
- DOI: 10.1093/toxsci/kfw089
Editor's Highlight: Hydroxyurea Exposure Activates the P53 Signaling Pathway in Murine Organogenesis-Stage Embryos
Abstract
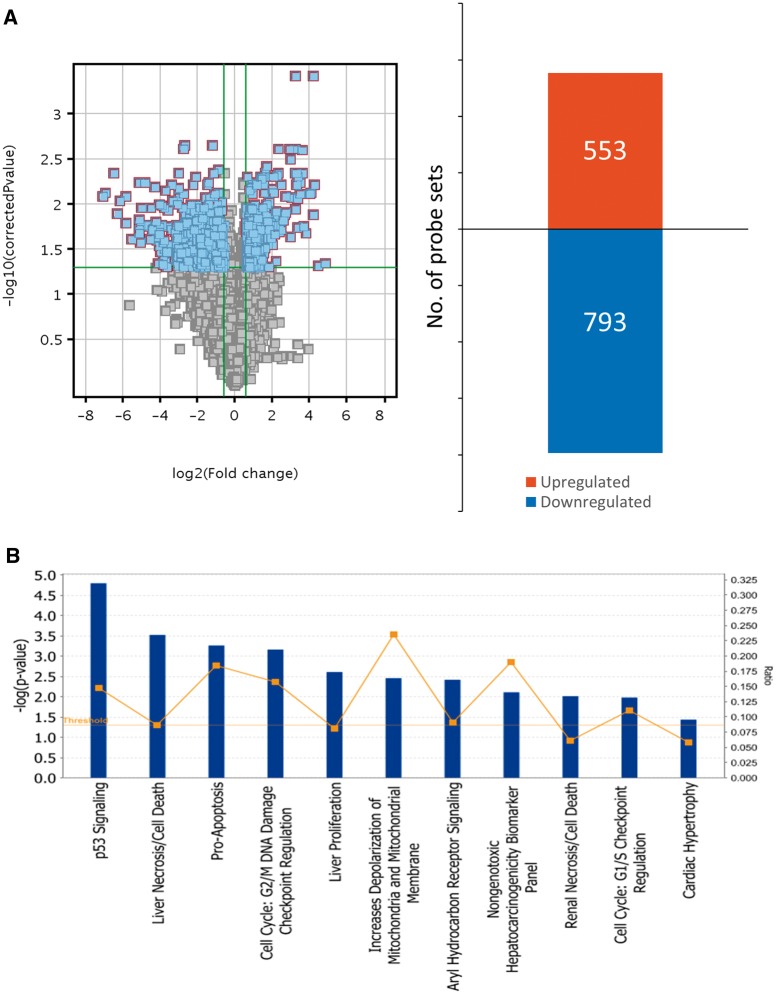
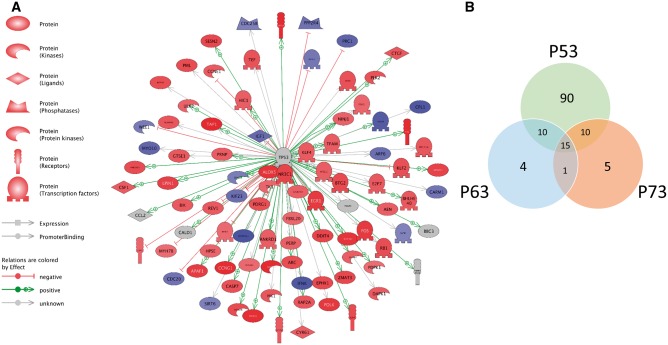
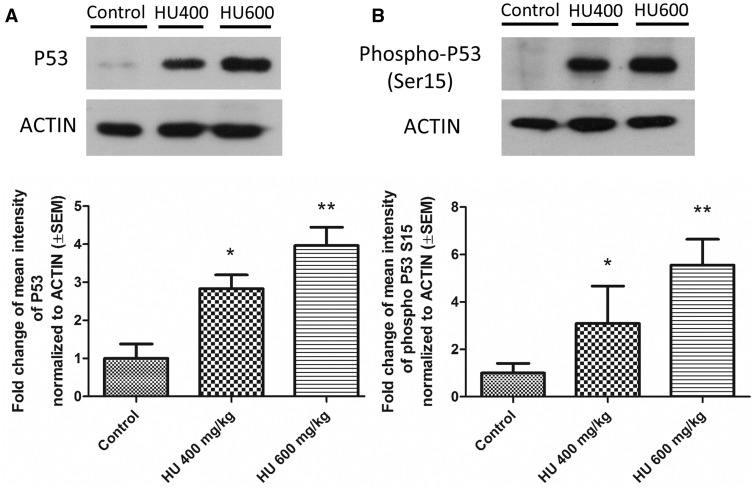
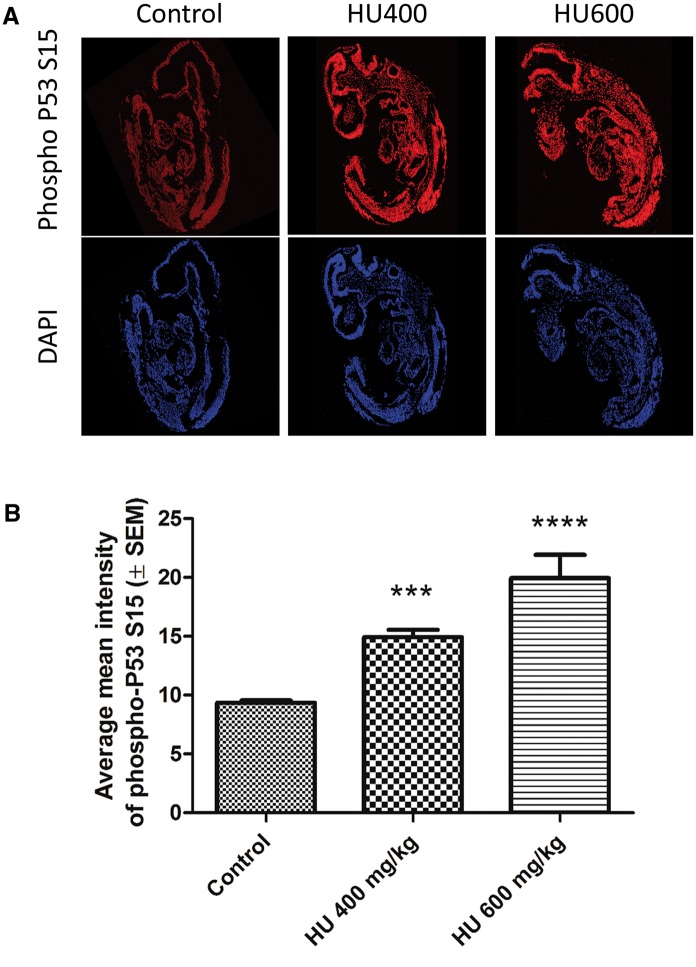
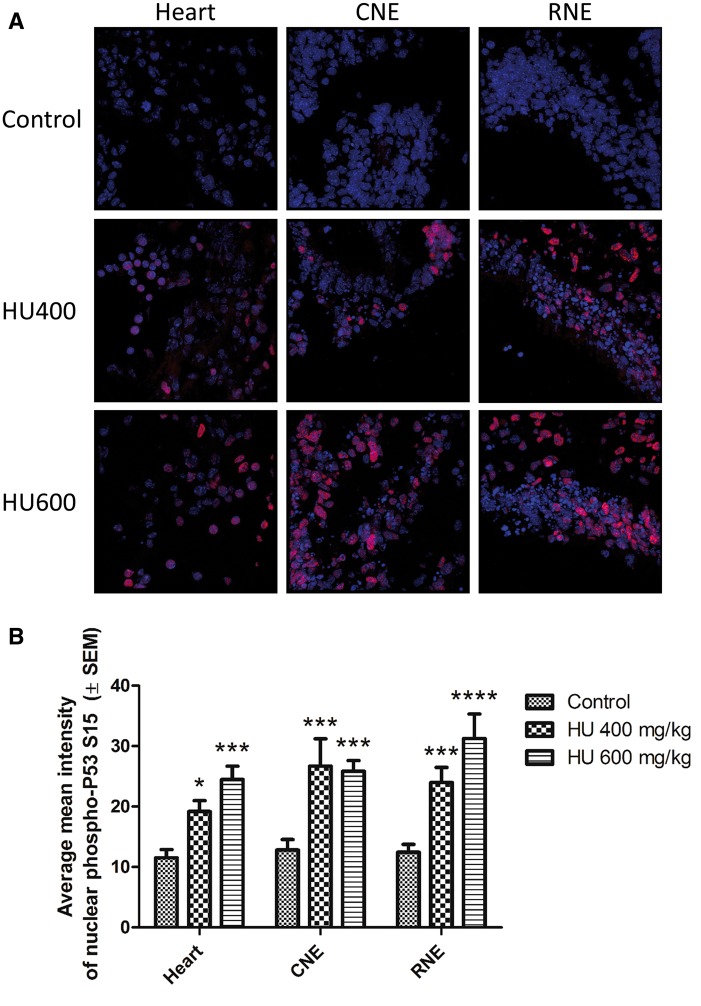
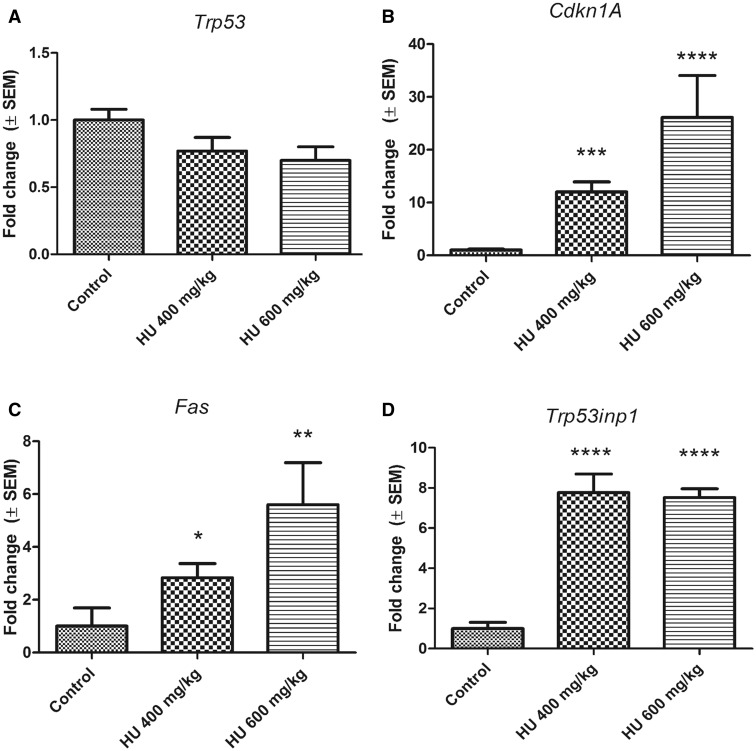
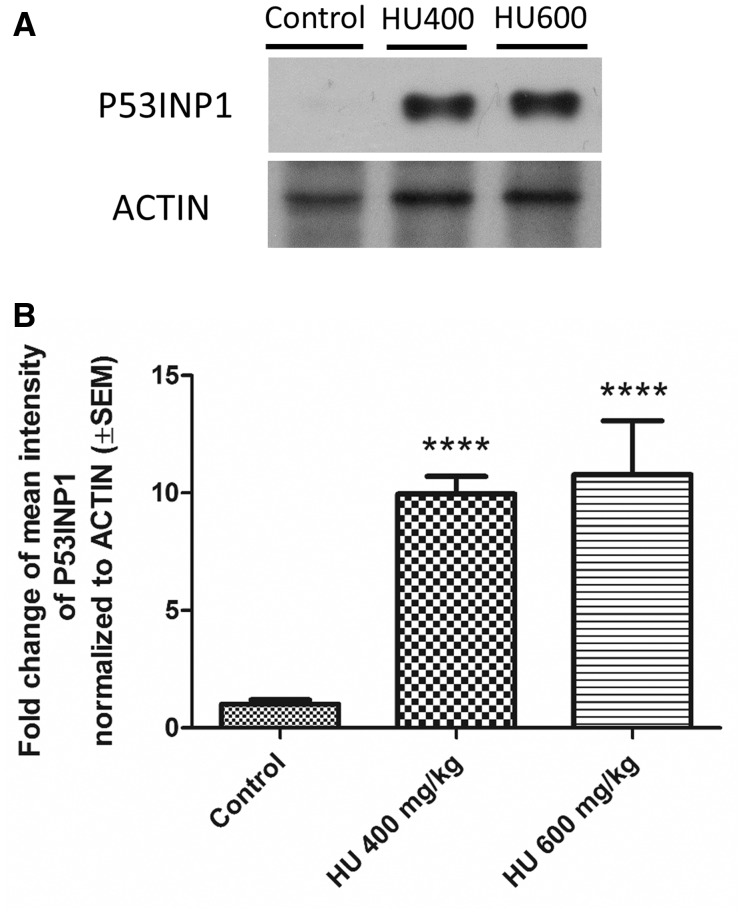
Hydroxyurea, an anticancer agent and potent teratogen, induces oxidative stress and activates a DNA damage response pathway in the gestation day (GD) 9 mouse embryo. To delineate the stress response pathways activated by this drug, we investigated the effect of hydroxyurea exposure on the transcriptome of GD 9 embryos. Timed pregnant CD-1 mice were treated with saline or hydroxyurea (400 mg/kg or 600 mg/kg) on GD 9; embryonic gene and protein expression were examined 3 h later. Microarray analysis revealed that the expression of 1346 probe sets changed significantly in embryos exposed to hydroxyurea compared with controls; the P53 signaling pathway was highly affected. In addition, P53 related family members, P63 and P73, were predicted to be activated and had common and unique downstream targets. Western blot analysis revealed that active phospho-P53 was significantly increased in drug-exposed embryos; confocal microscopy showed that the translocation of phospho-P53 to the nucleus was widespread in the embryo. Furthermore, qRT-PCR showed that the expression of P53-regulated genes (Cdkn1A, Fas, and Trp53inp1) was significantly upregulated in hydroxyurea-exposed embryos; the concentration of the redox sensitive P53INP1 protein was also increased in a hydroxyurea dose-dependent fashion. Thus, hydroxyurea elicits a significant effect on the transcriptome of the organogenesis stage murine embryo, activating several key developmental signaling pathways related to DNA damage and oxidative stress. We propose that the P53 pathway plays a central role in the embryonic stress response and the developmental outcome after teratogen exposure.
Keywords: birth defects; developmental toxicity; embryonic stress; teratogen.
© The Author 2016. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved. For Permissions, please e-mail: journals.permissions@oup.com.
Figures
Similar articles
-
The Roles of P53 and Its Family Proteins, P63 and P73, in the DNA Damage Stress Response in Organogenesis-Stage Mouse Embryos.Toxicol Sci. 2018 Apr 1;162(2):439-449. doi: 10.1093/toxsci/kfx270. Toxicol Sci. 2018. PMID: 29228353 Free PMC article.
-
Hydroxyurea embryotoxicity is enhanced in P53-deficient mice.Reprod Toxicol. 2018 Oct;81:28-33. doi: 10.1016/j.reprotox.2018.06.011. Epub 2018 Jun 22. Reprod Toxicol. 2018. PMID: 29940331
-
Deprenyl enhances the teratogenicity of hydroxyurea in organogenesis stage mouse embryos.Toxicol Sci. 2013 Aug;134(2):391-9. doi: 10.1093/toxsci/kft115. Epub 2013 May 21. Toxicol Sci. 2013. PMID: 23696560 Free PMC article.
-
Mechanisms of the embryo's response to embryopathic stressors: a focus on p53.J Reprod Immunol. 2010 May;85(1):76-80. doi: 10.1016/j.jri.2010.01.003. Epub 2010 Mar 12. J Reprod Immunol. 2010. PMID: 20227113 Review.
-
Involvement of p53-dependent apoptosis in radiation teratogenesis and in the radioadaptive response in the late organogenesis of mice.J Radiat Res. 2001 Mar;42(1):1-10. doi: 10.1269/jrr.42.1. J Radiat Res. 2001. PMID: 11393885 Review.
Cited by
-
Redox mechanisms of environmental toxicants on male reproductive function.Front Cell Dev Biol. 2024 Feb 26;12:1333845. doi: 10.3389/fcell.2024.1333845. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 38469179 Free PMC article. Review.
-
The Roles of P53 and Its Family Proteins, P63 and P73, in the DNA Damage Stress Response in Organogenesis-Stage Mouse Embryos.Toxicol Sci. 2018 Apr 1;162(2):439-449. doi: 10.1093/toxsci/kfx270. Toxicol Sci. 2018. PMID: 29228353 Free PMC article.
-
Suppression of Trim32 Enhances Motor Function Repair after Traumatic Brain Injury Associated with Antiapoptosis.Cell Transplant. 2017 Jul;26(7):1276-1285. doi: 10.1177/0963689717716510. Cell Transplant. 2017. PMID: 28933219 Free PMC article.
-
Hydroxyurea affects in vitro porcine oocyte maturation through increased apoptosis and oxidative stress.Biosci Rep. 2021 Apr 30;41(4):BSR20203091. doi: 10.1042/BSR20203091. Biosci Rep. 2021. PMID: 33844009 Free PMC article.
-
The Redox Theory of Development.Antioxid Redox Signal. 2020 Apr 1;32(10):715-740. doi: 10.1089/ars.2019.7976. Antioxid Redox Signal. 2020. PMID: 31891515 Free PMC article. Review.
References
-
- Adachi S., Cross A. R., Babior B. M., Gottlieb R. A. (1997). Bcl-2 and the outer mitochondrial membrane in the inactivation of cytochrome c during Fas-mediated apoptosis. J. Biol. Chem. 272, 21878–21882. - PubMed
-
- Armstrong J. F., Kaufman M. H., Harrison D. J., Clarke A. R. (1995). High-frequency developmental abnormalities in p53-deficient mice. Curr. Biol. 5, 931–936. - PubMed
-
- Aylon Y., Oren M. (2007). Living with p53, dying of p53. Cell 130, 597–600. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
