

Combinatorial effects of odorants on mouse behavior
- PMID: 27208093
- PMCID: PMC4988607
- DOI: 10.1073/pnas.1605973113
Combinatorial effects of odorants on mouse behavior
Abstract
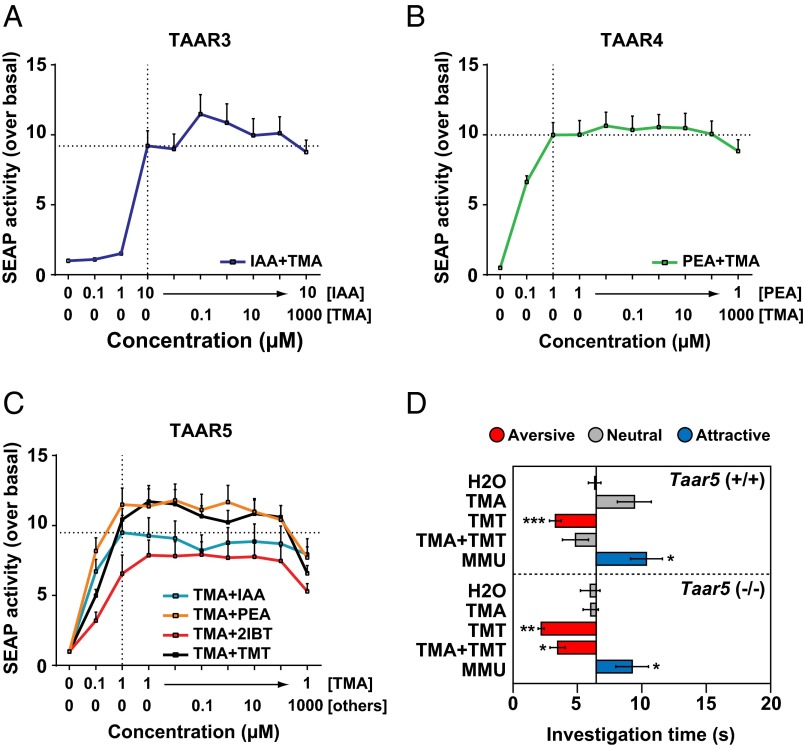
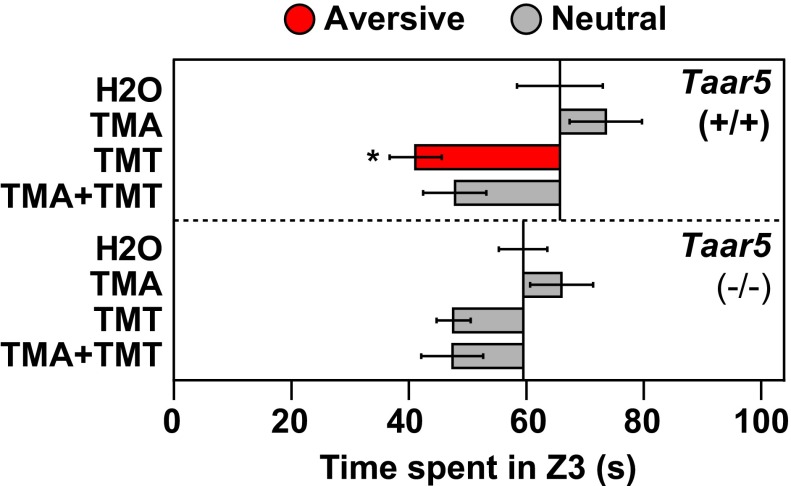
The mechanisms by which odors induce instinctive behaviors are largely unknown. Odor detection in the mouse nose is mediated by >1, 000 different odorant receptors (ORs) and trace amine-associated receptors (TAARs). Odor perceptions are encoded combinatorially by ORs and can be altered by slight changes in the combination of activated receptors. However, the stereotyped nature of instinctive odor responses suggests the involvement of specific receptors and genetically programmed neural circuits relatively immune to extraneous odor stimuli and receptor inputs. Here, we report that, contrary to expectation, innate odor-induced behaviors can be context-dependent. First, different ligands for a given TAAR can vary in behavioral effect. Second, when combined, some attractive and aversive odorants neutralize one another's behavioral effects. Both a TAAR ligand and a common odorant block aversion to a predator odor, indicating that this ability is not unique to TAARs and can extend to an aversive response of potential importance to survival. In vitro testing of single receptors with binary odorant mixtures indicates that behavioral blocking can occur without receptor antagonism in the nose. Moreover, genetic ablation of a single receptor prevents its cognate ligand from blocking predator odor aversion, indicating that the blocking requires sensory input from the receptor. Together, these findings indicate that innate odor-induced behaviors can depend on context, that signals from a single receptor can block innate odor aversion, and that instinctive behavioral responses to odors can be modulated by interactions in the brain among signals derived from different receptors.
Keywords: TAAR; behavior; odorants; olfaction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
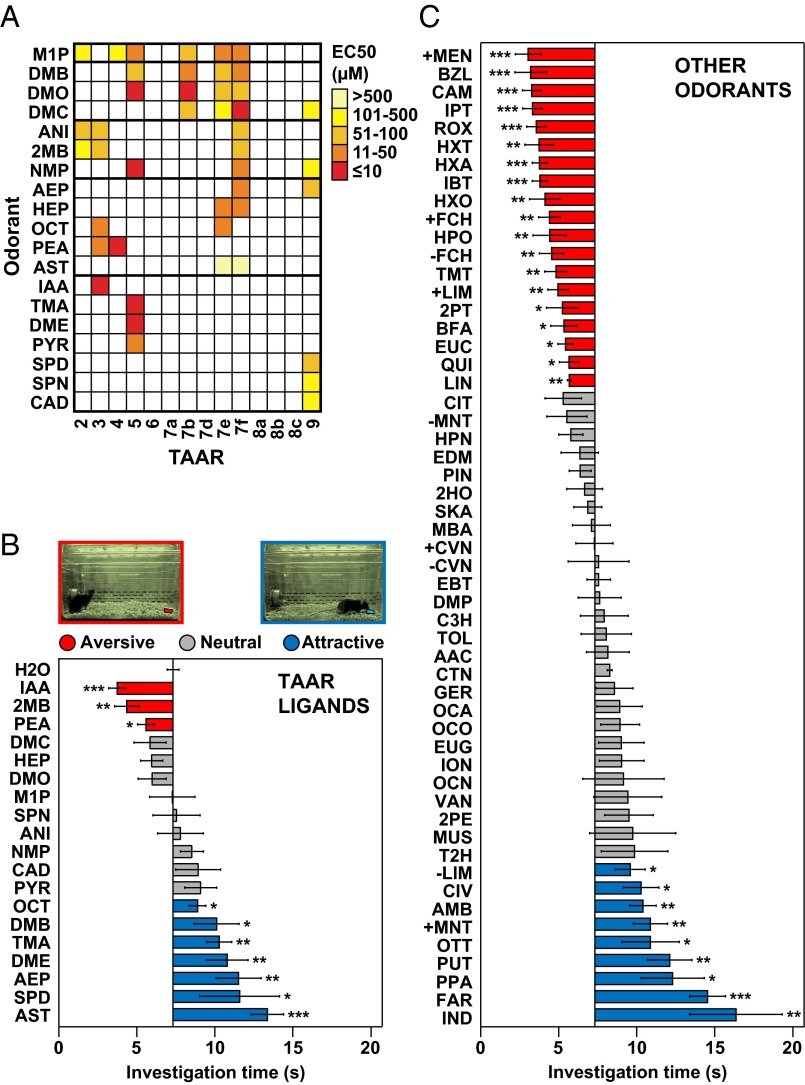
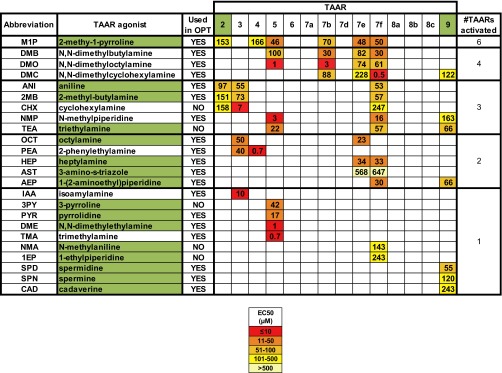
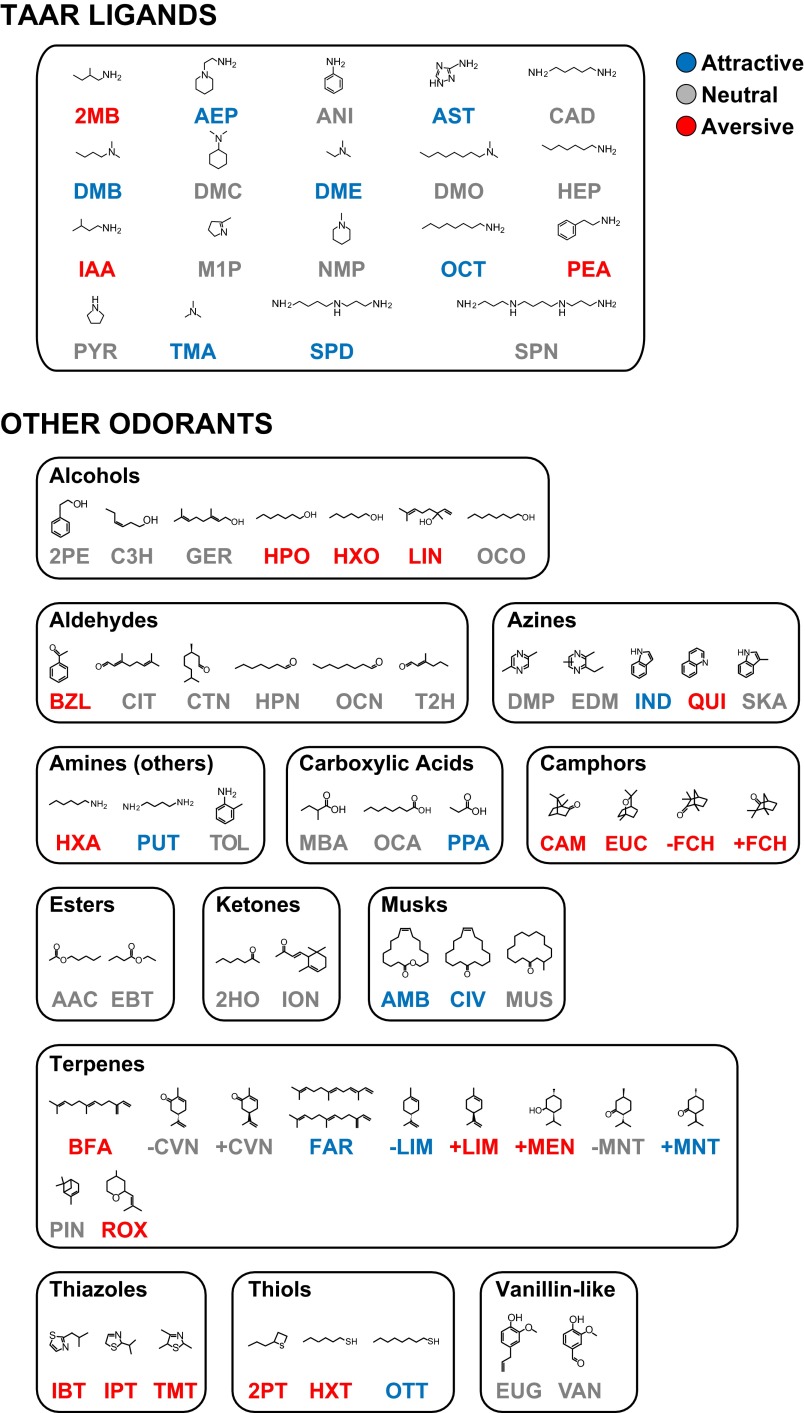
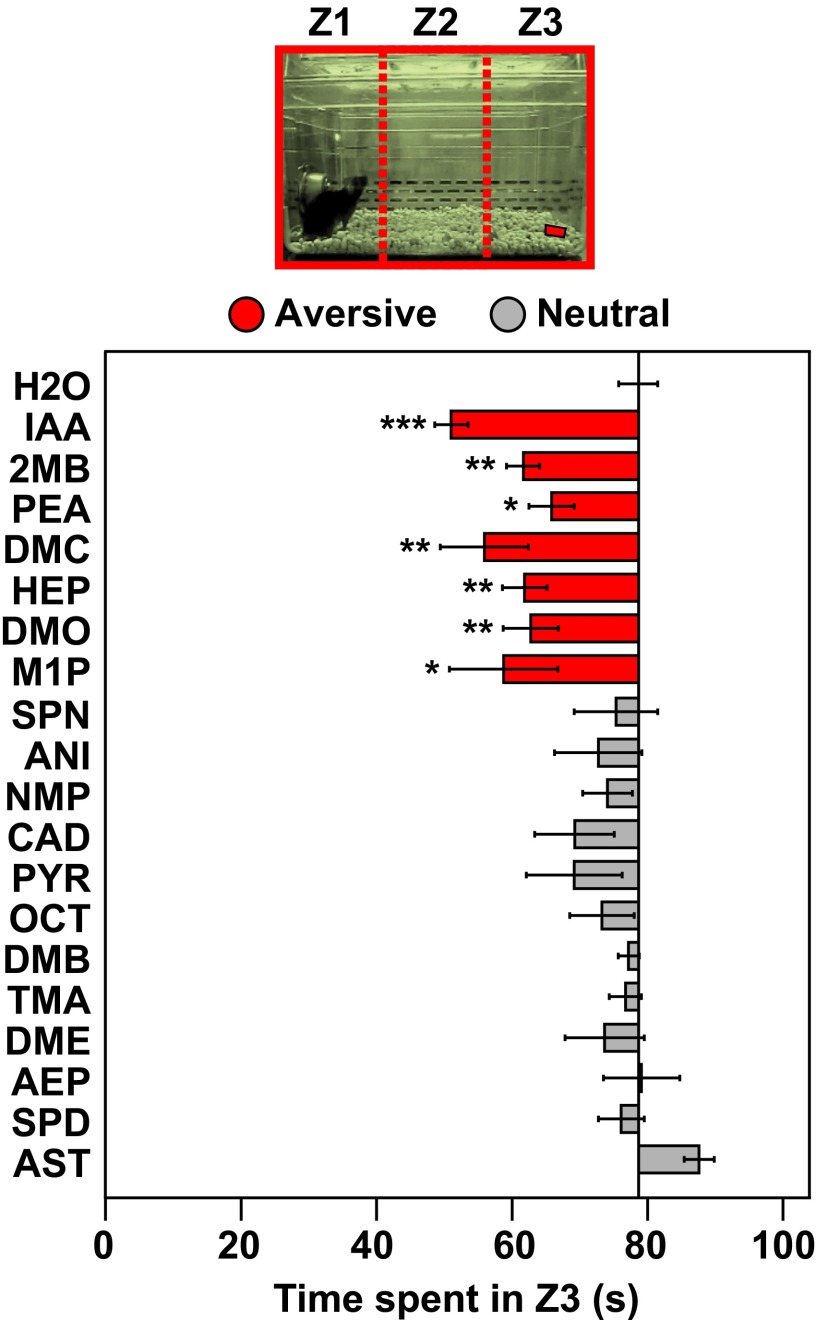
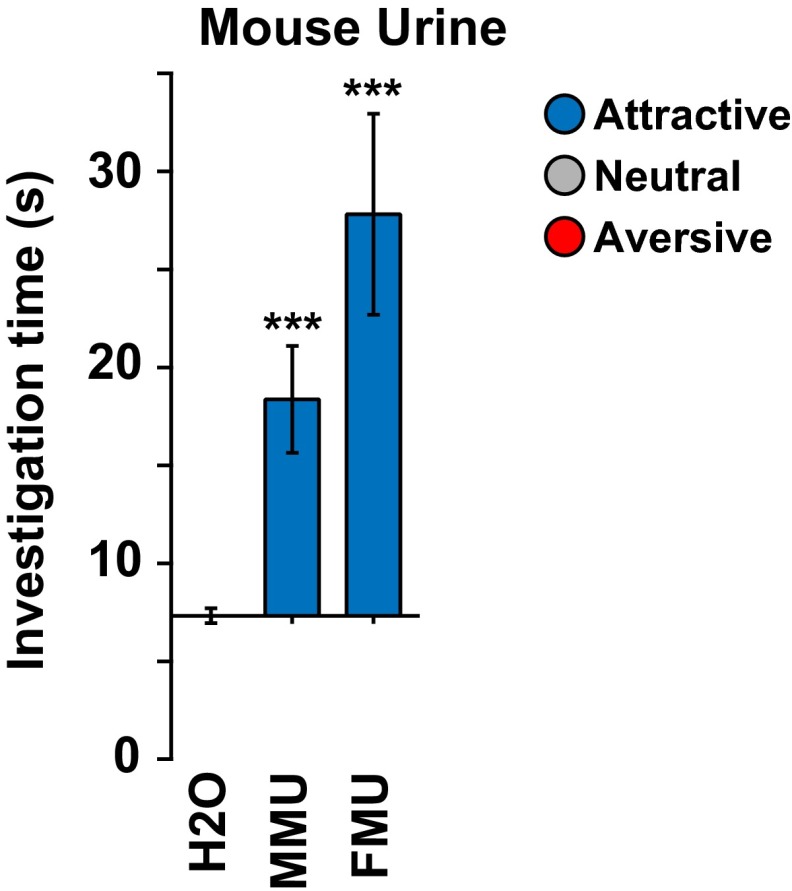
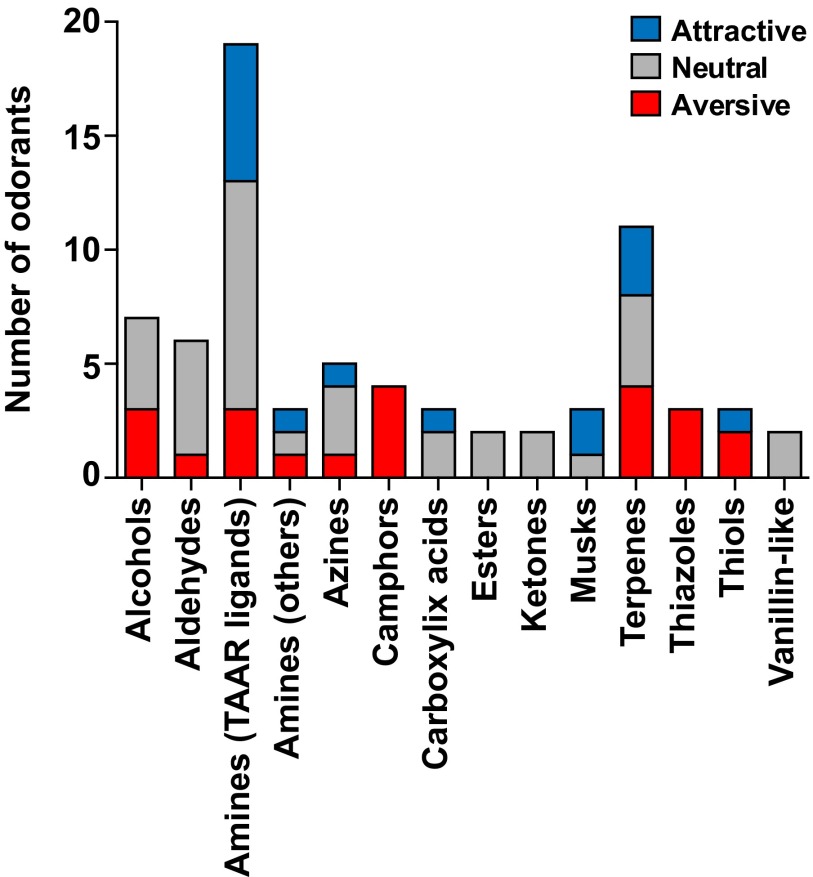
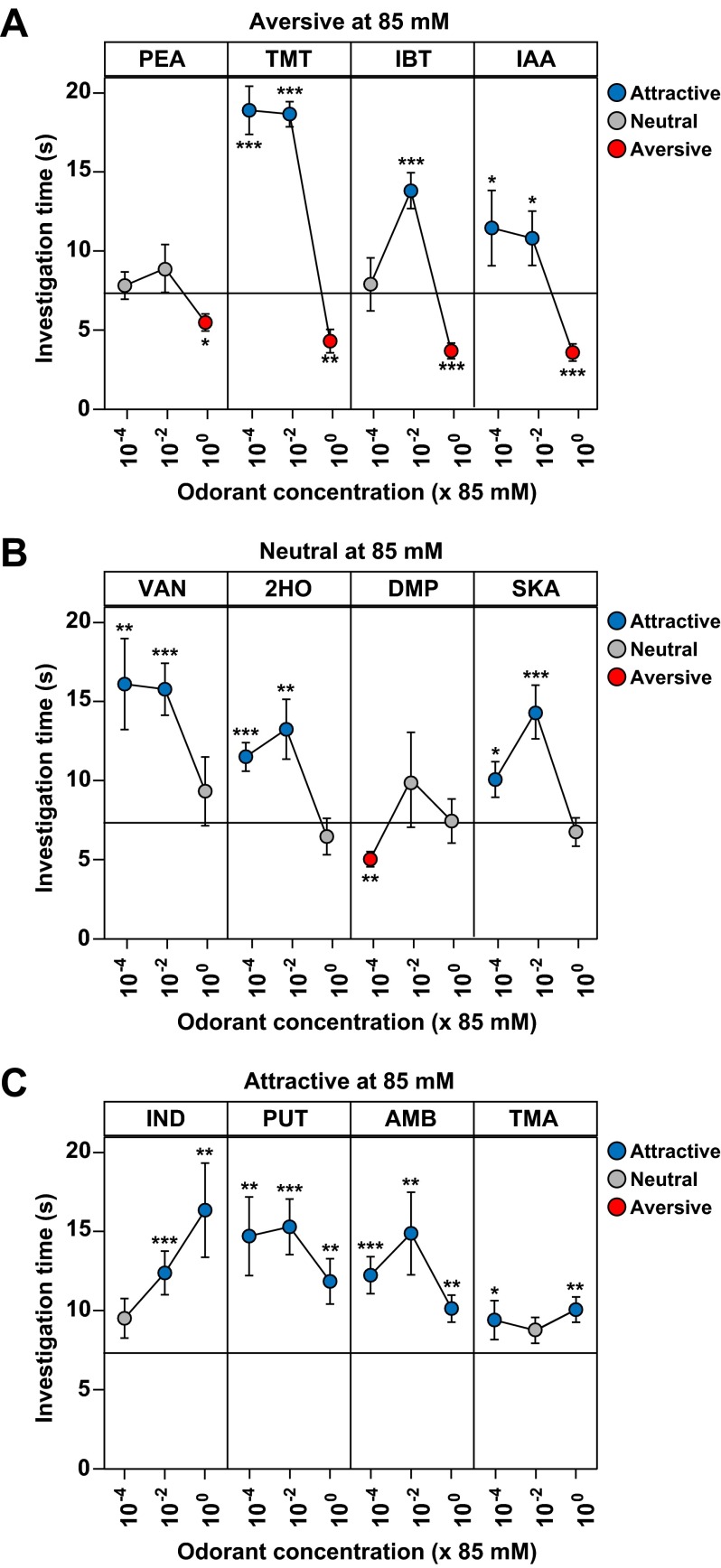
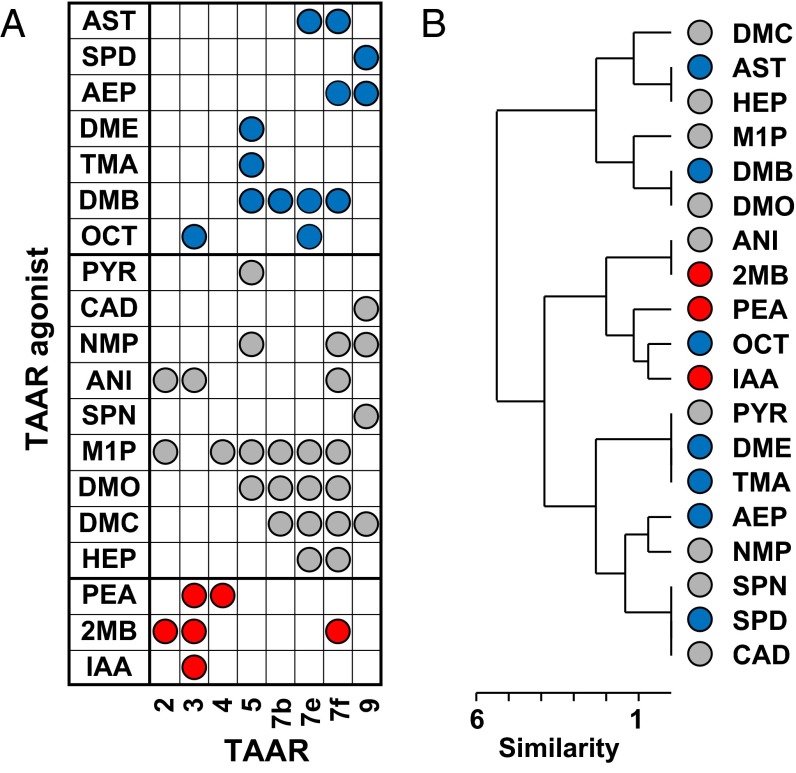
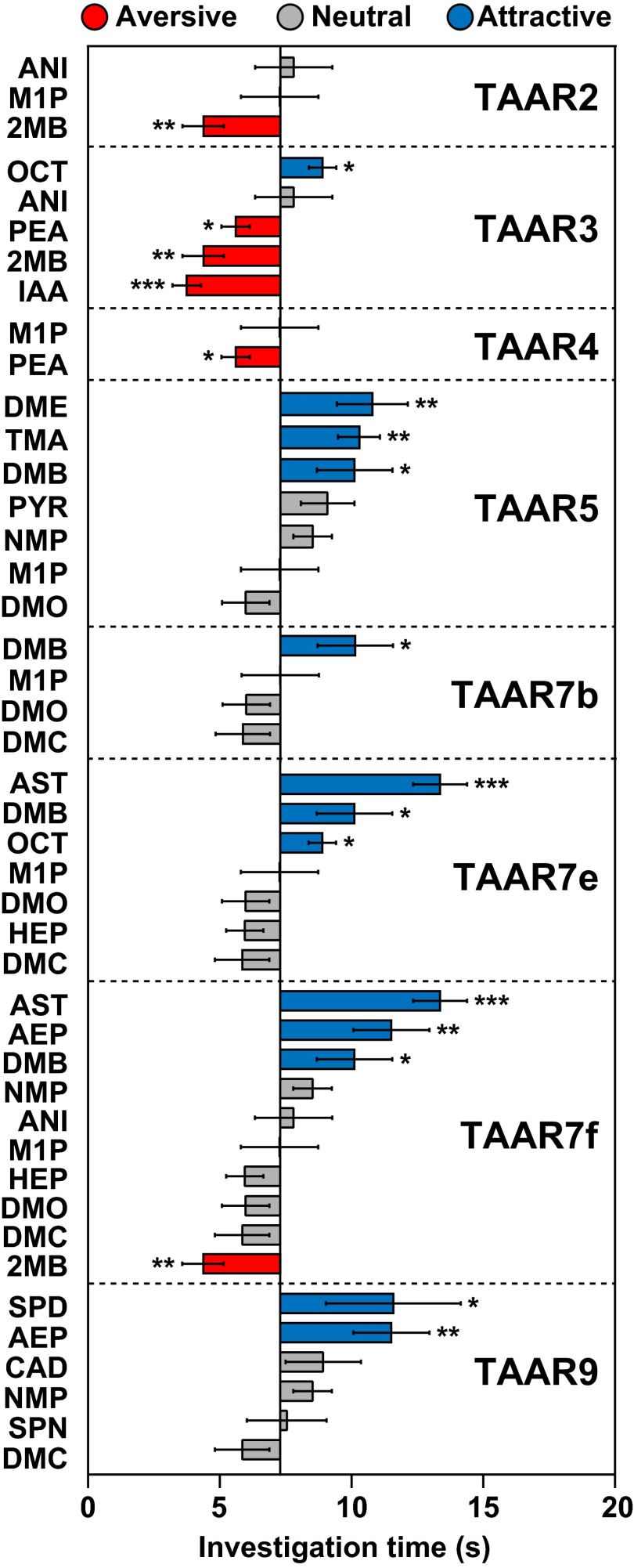
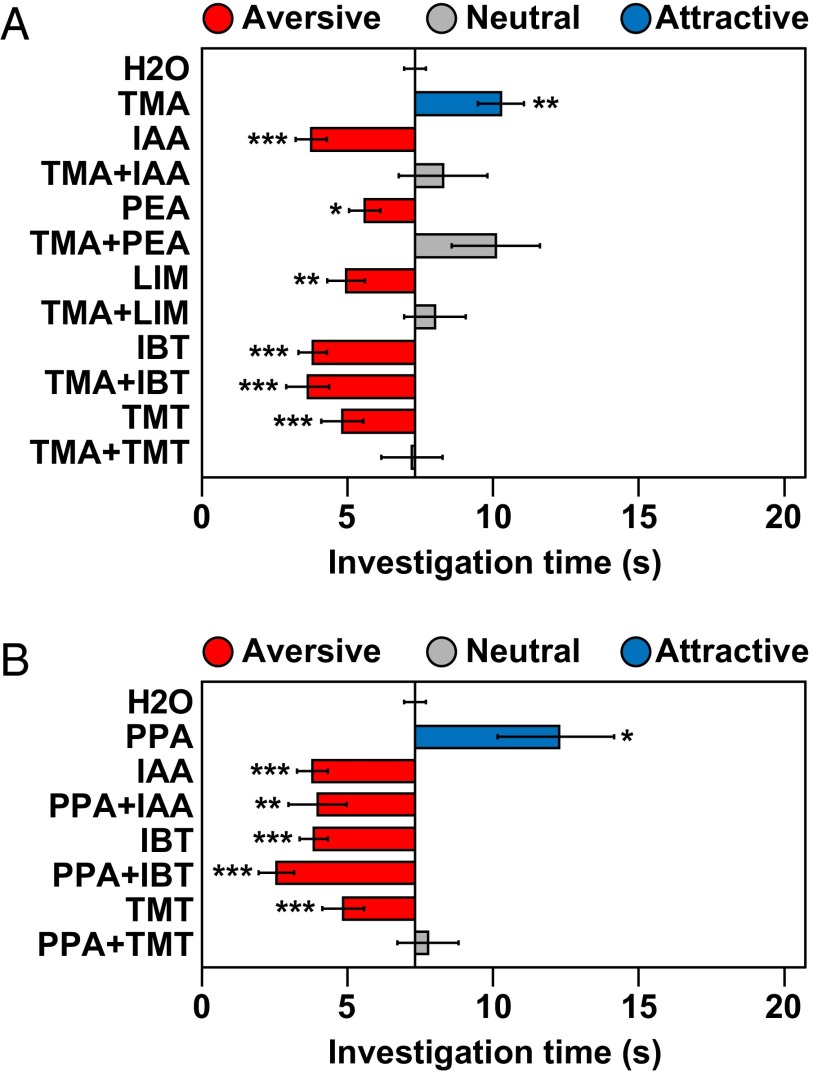
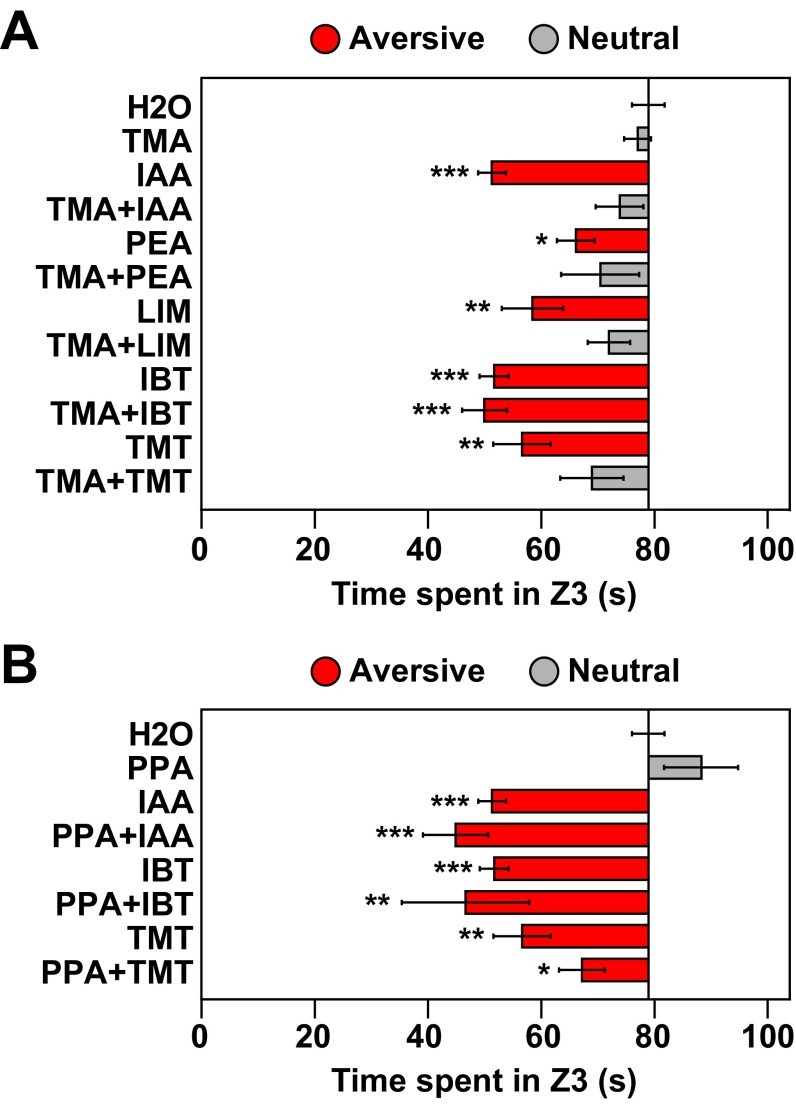
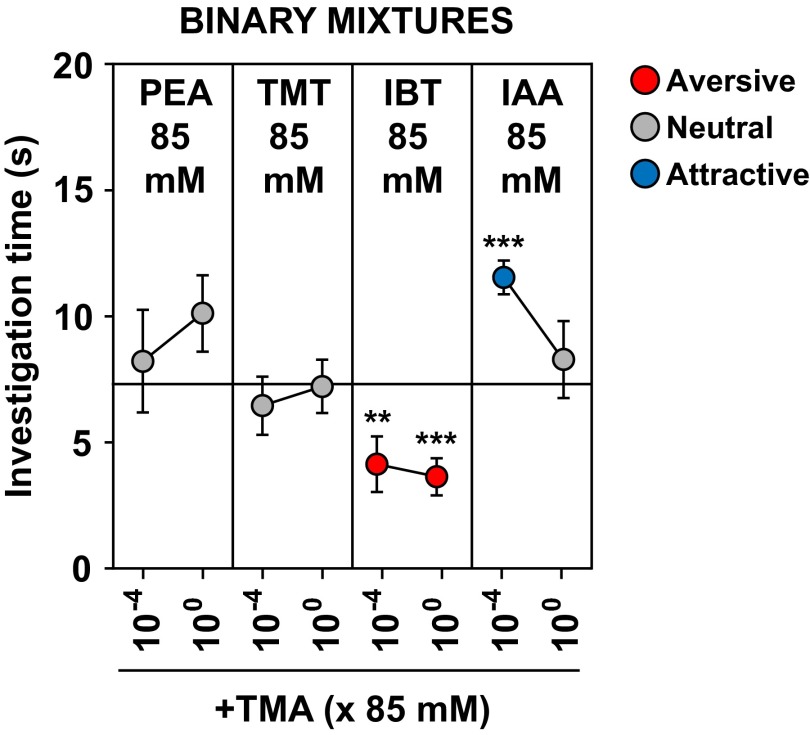
References
-
- Buck LB, Bargmann C. Smell and taste: The chemical senses. In: Kandel E, Schwartz J, Jessell T, Siegelbaum S, Hudspeth AJ, editors. Principles of Neuroscience. McGraw-Hill; New York: 2012. pp. 712–742.
-
- Munger SD, Leinders-Zufall T, Zufall F. Subsystem organization of the mammalian sense of smell. Annu Rev Physiol. 2009;71:115–140. - PubMed
-
- Buck L, Axel R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell. 1991;65(1):175–187. - PubMed
-
- Ressler KJ, Sullivan SL, Buck LB. A zonal organization of odorant receptor gene expression in the olfactory epithelium. Cell. 1993;73(3):597–609. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
