

Membrane-Induced Folding of the Plant Stress Dehydrin Lti30
- PMID: 27208263
- PMCID: PMC4902575
- DOI: 10.1104/pp.15.01531
Membrane-Induced Folding of the Plant Stress Dehydrin Lti30
Abstract
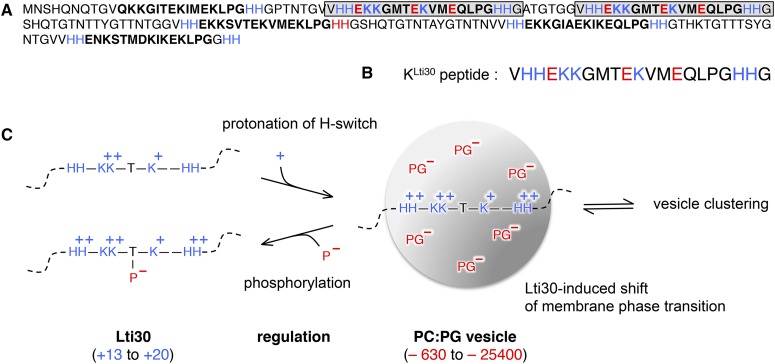
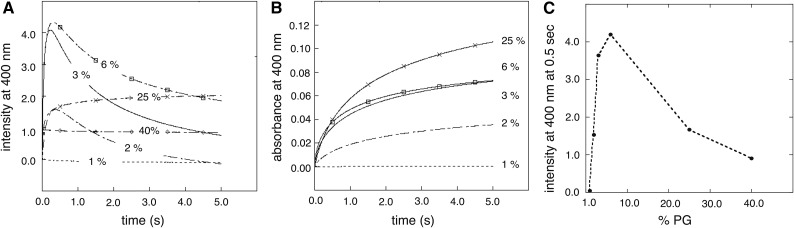
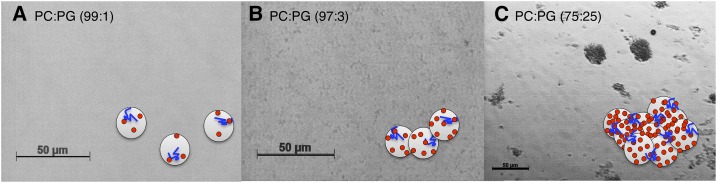
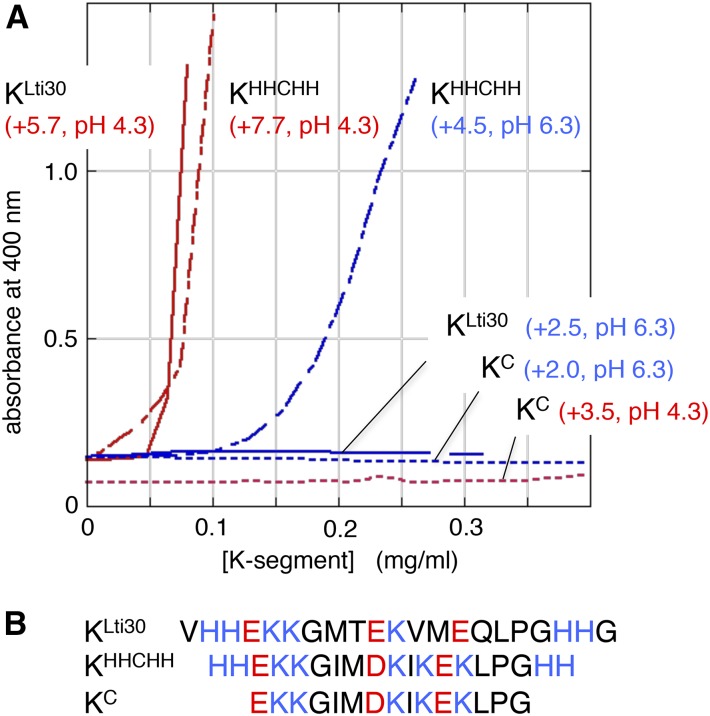
Dehydrins are disordered proteins that are expressed in plants as a response to embryogenesis and water-related stress. The molecular function and structural action of the dehydrins are yet elusive, but increasing evidence points to a role in protecting the structure and functional dynamics of cell membranes. An intriguing example is the cold-induced dehydrin Lti30 that binds to membranes by its conserved K segments. Moreover, this binding can be regulated by pH and phosphorylation and shifts the membrane phase transition to lower temperatures, consistent with the protein's postulated function in cold stress. In this study, we reveal how the Lti30-membrane interplay works structurally at atomic level resolution in Arabidopsis (Arabidopsis thaliana). Nuclear magnetic resonance analysis suggests that negatively charged lipid head groups electrostatically capture the protein's disordered K segments, which locally fold up into α-helical segments on the membrane surface. Thus, Lti30 conforms to the general theme of structure-function relationships by folding upon binding, in spite of its disordered, atypically hydrophilic and repetitive sequence signatures. Moreover, the fixed and well-defined structure of the membrane-bound K segments suggests that dehydrins have the molecular prerequisites for higher level binding specificity and regulation, raising new questions about the complexity of their biological function.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures
Similar articles
-
Tunable membrane binding of the intrinsically disordered dehydrin Lti30, a cold-induced plant stress protein.Plant Cell. 2011 Jun;23(6):2391-404. doi: 10.1105/tpc.111.085183. Epub 2011 Jun 10. Plant Cell. 2011. PMID: 21665998 Free PMC article.
-
The disordered plant dehydrin Lti30 protects the membrane during water-related stress by cross-linking lipids.J Biol Chem. 2019 Apr 19;294(16):6468-6482. doi: 10.1074/jbc.RA118.007163. Epub 2019 Feb 28. J Biol Chem. 2019. PMID: 30819802 Free PMC article.
-
Cryoprotective mechanism of a small intrinsically disordered dehydrin protein.Protein Sci. 2011 Jan;20(1):42-50. doi: 10.1002/pro.534. Protein Sci. 2011. PMID: 21031484 Free PMC article.
-
Plant aquaporins: novel functions and regulation properties.FEBS Lett. 2007 May 25;581(12):2227-36. doi: 10.1016/j.febslet.2007.03.021. Epub 2007 Mar 15. FEBS Lett. 2007. PMID: 17382935 Review.
-
Structural and Functional Dynamics of Dehydrins: A Plant Protector Protein under Abiotic Stress.Int J Mol Sci. 2018 Oct 31;19(11):3420. doi: 10.3390/ijms19113420. Int J Mol Sci. 2018. PMID: 30384475 Free PMC article. Review.
Cited by
-
Multiple copies of a novel amphipathic α-helix forming segment in Physcomitrella patens dehydrin play a key role in abiotic stress mitigation.J Biol Chem. 2021 Jan-Jun;296:100596. doi: 10.1016/j.jbc.2021.100596. Epub 2021 Mar 26. J Biol Chem. 2021. PMID: 33781743 Free PMC article.
-
Functional Characteristics Analysis of Dehydrins in Larix kaempferi under Osmotic Stress.Int J Mol Sci. 2021 Feb 9;22(4):1715. doi: 10.3390/ijms22041715. Int J Mol Sci. 2021. PMID: 33572055 Free PMC article.
-
Dehydrin CaDHN2 Enhances Drought Tolerance by Affecting Ascorbic Acid Synthesis under Drought in Peppers.Plants (Basel). 2023 Nov 18;12(22):3895. doi: 10.3390/plants12223895. Plants (Basel). 2023. PMID: 38005792 Free PMC article.
-
The N-Terminal Region of Soybean PM1 Protein Protects Liposomes during Freeze-Thaw.Int J Mol Sci. 2020 Aug 3;21(15):5552. doi: 10.3390/ijms21155552. Int J Mol Sci. 2020. PMID: 32756462 Free PMC article.
-
The Halophyte Dehydrin Sequence Landscape.Biomolecules. 2022 Feb 19;12(2):330. doi: 10.3390/biom12020330. Biomolecules. 2022. PMID: 35204830 Free PMC article.
References
-
- Alsheikh MK, Heyen BJ, Randall SK (2003) Ion binding properties of the dehydrin ERD14 are dependent upon phosphorylation. J Biol Chem 278: 40882–40889 - PubMed
-
- Alsheikh MK, Svensson JT, Randall SK (2005) Phosphorylation regulated ion-binding is a property shared by the acidic subclass dehydrins. Plant Cell Environ 28: 1114–1122
-
- Andersson A, Mäler L (2005) Magnetic resonance investigations of lipid motion in isotropic bicelles. Langmuir 21: 7702–7709 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
