

Corpse Engulfment Generates a Molecular Memory that Primes the Macrophage Inflammatory Response
- PMID: 27212238
- PMCID: PMC4912690
- DOI: 10.1016/j.cell.2016.04.049
Corpse Engulfment Generates a Molecular Memory that Primes the Macrophage Inflammatory Response
Abstract
Macrophages are multifunctional cells that perform diverse roles in health and disease. Emerging evidence has suggested that these innate immune cells might also be capable of developing immunological memory, a trait previously associated with the adaptive system alone. While recent studies have focused on the dramatic macrophage reprogramming that follows infection and protects against secondary microbial attack, can macrophages also develop memory in response to other cues? Here, we show that apoptotic corpse engulfment by Drosophila macrophages is an essential primer for their inflammatory response to tissue damage and infection in vivo. Priming is triggered via calcium-induced JNK signaling, which leads to upregulation of the damage receptor Draper, thus providing a molecular memory that allows the cell to rapidly respond to subsequent injury or infection. This remarkable plasticity and capacity for memory places macrophages as key therapeutic targets for treatment of inflammatory disorders.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures



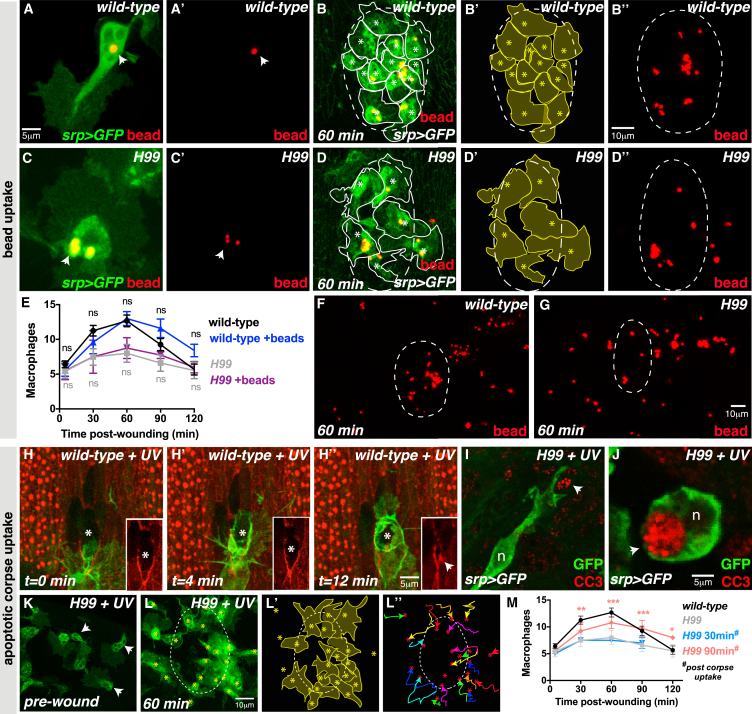


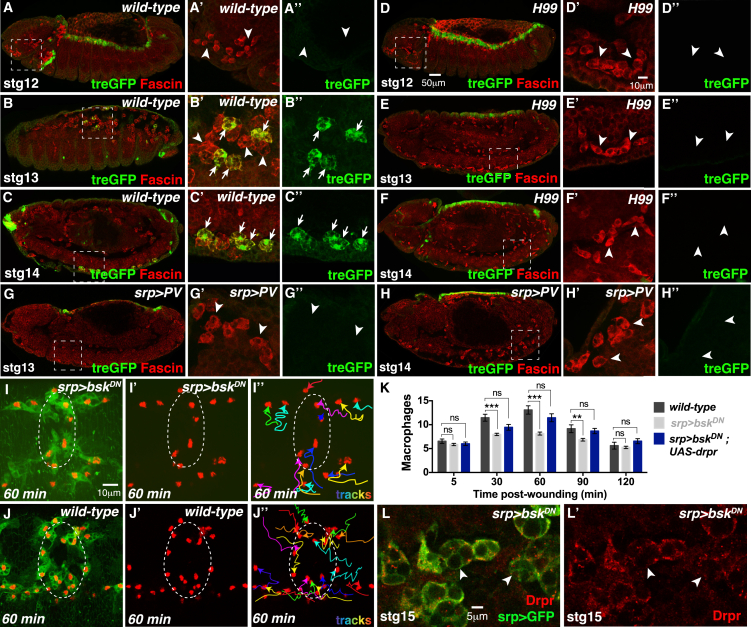

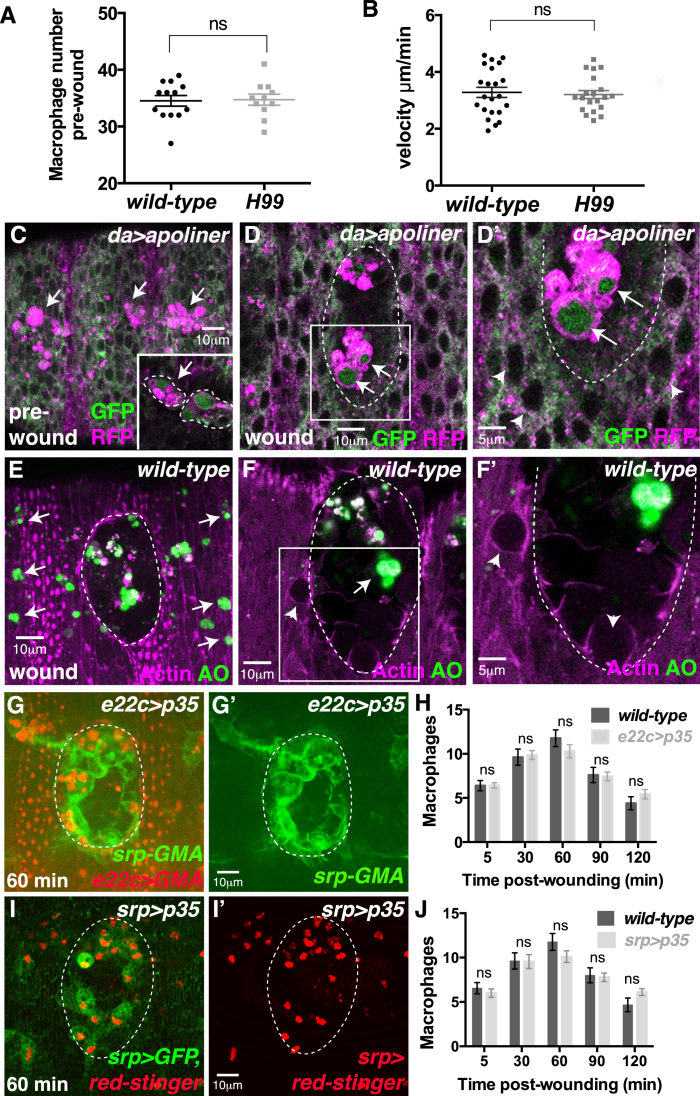



Comment in
-
Innate Immune Memory: Activation of Macrophage Killing Ability by Developmental Duties.Curr Biol. 2016 Jun 20;26(12):R503-R505. doi: 10.1016/j.cub.2016.05.016. Curr Biol. 2016. PMID: 27326712
-
Macrophages: Memories of the dead give strength.Nat Rev Immunol. 2016 Aug;16(8):464-5. doi: 10.1038/nri.2016.81. Epub 2016 Jul 4. Nat Rev Immunol. 2016. PMID: 27374638 No abstract available.
References
-
- Adachi-Yamada T., Nakamura M., Irie K., Tomoyasu Y., Sano Y., Mori E., Goto S., Ueno N., Nishida Y., Matsumoto K. p38 mitogen-activated protein kinase can be involved in transforming growth factor beta superfamily signal transduction in Drosophila wing morphogenesis. Mol. Cell. Biol. 1999;19:2322–2329. - PMC - PubMed
-
- Bistoni F., Verducci G., Perito S., Vecchiarelli A., Puccetti P., Marconi P., Cassone A. Immunomodulation by a low-virulence, agerminative variant of Candida albicans. Further evidence for macrophage activation as one of the effector mechanisms of nonspecific anti-infectious protection. J. Med. Vet. Mycol. 1988;26:285–299. - PubMed
-
- Blok B.A., Arts R.J.W., van Crevel R., Benn C.S., Netea M.G. Trained innate immunity as underlying mechanism for the long-term, nonspecific effects of vaccines. J. Leukoc. Biol. 2015;98:347–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
