

Review
doi: 10.1007/s00439-016-1683-5.
Epub 2016 May 23.
The hnRNP family: insights into their role in health and disease
Affiliations
- PMID: 27215579
- PMCID: PMC4947485
- DOI: 10.1007/s00439-016-1683-5
Item in Clipboard
Review
The hnRNP family: insights into their role in health and disease
Hum Genet.
2016 Aug.
Abstract
Heterogeneous nuclear ribonucleoproteins (hnRNPs) represent a large family of RNA-binding proteins (RBPs) that contribute to multiple aspects of nucleic acid metabolism including alternative splicing, mRNA stabilization, and transcriptional and translational regulation. Many hnRNPs share general features, but differ in domain composition and functional properties. This review will discuss the current knowledge about the different hnRNP family members, focusing on their structural and functional divergence. Additionally, we will highlight their involvement in neurodegenerative diseases and cancer, and the potential to develop RNA-based therapies.
Figures
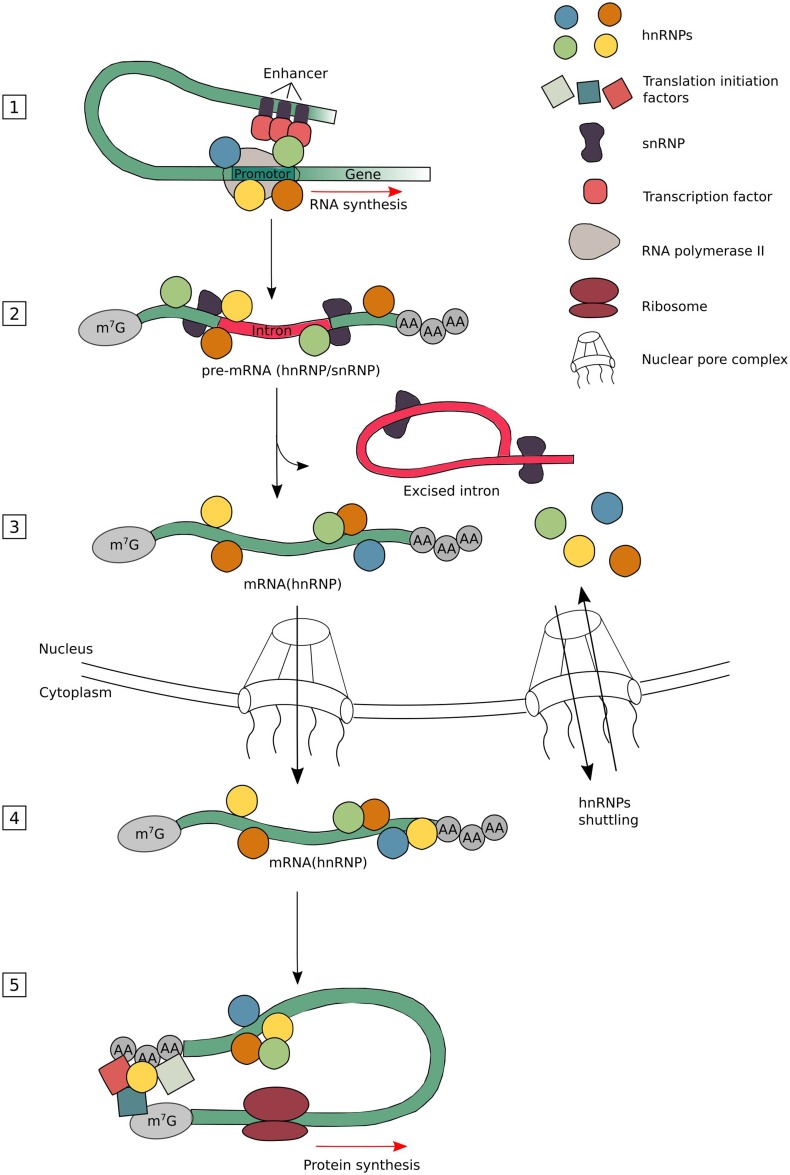
The diverse nuclear and cytoplasmic functions of hnRNPs. The hnRNPs have been found to be involved in different stages of the mRNA metabolism. They constantly undergo a binding and release from their target mRNAs depending on the modification needed. The hnRNPs bind, together with several transcription factors and other RBPs, to promotor and enhancer sequences to direct transcription (1). As soon as a part of the newly formed transcript is released by the RNA polymerase II, the hnRNPs and snRNPs bind rapidly to their nascent transcript to stabilize it (2). Once the correct RBP complexes are formed, intronic sequences are removed by the spliceosome. Many hnRNPs are known to regulate alternative splicing leading to exon skipping or intron retention (3). Mature mRNAs are stabilized by the binding of several types of RBPs, including hnRNPs, which are found necessary for export through the nuclear pore complex (NPC) and its transport through the cytoplasm until translation initiates (4). As hnRNPs have the capacity to bind to the 3′- and 5′-UTRs of mRNAs, they can control translational repression or enhancement. Depending on the composition of complexes at the sites where translation initiation factors assemble, the choice to start translation or not will be made (5). Please note that not all different hnRNP subgroup members are involved in every nuclear or cytoplasmic function. The different hnRNPs are color coded, but do not represent specific hnRNP members
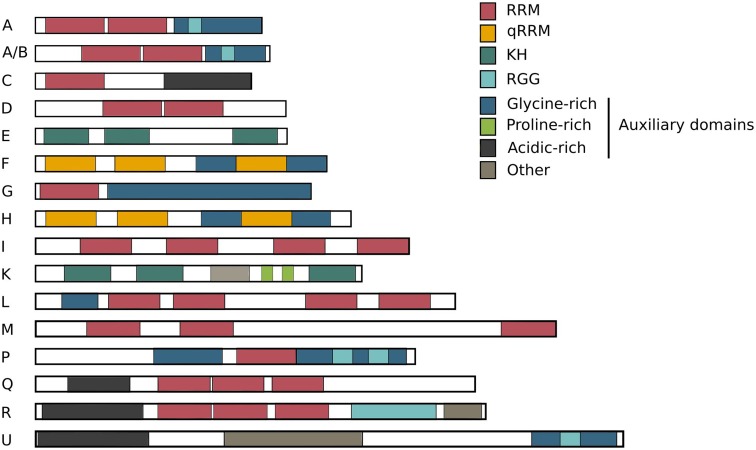
The hnRNP family. The hnRNPs have different molecular weights ranging from 34 to 120 kDa and are named alphabetically from hnRNP A1 to hnRNP U. Many hnRNPs are found to be present in the same complexes, all suggesting that multiple hnRNPs share a common structure and function. As shown in the overview, several structural domains are shared between different family members. The members of the hnRNP family are built up of four unique RNA-binding domains (RBDs). It is obvious that multiple family members carry the same RBDs, partly explaining their shared RNA-binding properties: RRM RNA recognition motif, qRRM quasi-RNA recognition motif and KH K-homology domain, RGG RNA-binding domain consisting of Arg-Gly-Gly repeats. The sizes of these 16 common hnRNPs are drawn relative to each other
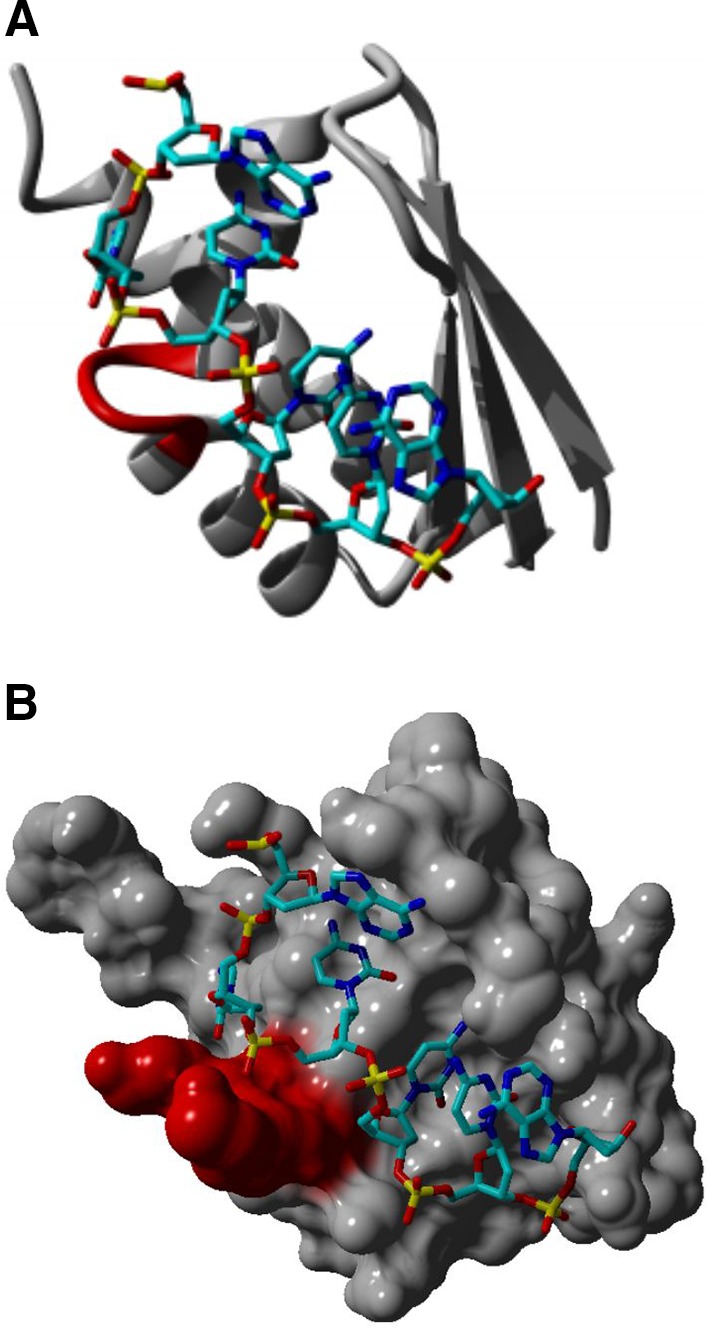
KH–RNA interaction. The KH domain was identified in multiple RNA-binding proteins and characterized by a 45 amino acid repeat that can be split into two groups. The Type I KH domains have a βα extension in their C-terminus, whereas the Type II KH domains have an αβ extension in their N-terminus. The core region of the KH domain is characterized by three-stranded antiparallel β-sheets together with three α-helices (βααββα). It is believed that the nucleotide recognition of the KH domain is determined by a conserved GxxG loop (highlighted in red) in the nucleotide stretch that links two α-helices in the KH core (a). This leads to the orientation of four nucleotides toward the groove in the protein structure where the nucleotide (backbone colored in light and dark blue) recognition is mainly determined by hydrophobic interactions and hydrogen bonds. Besides sequence-specific recognition, the overall shape of the KH hydrophobic groove, which is determined by the conformation of multiple side chains, is equally important in KH–RNA recognition (b). It was shown by using surface plasmon resonance that the KH domain shows a high affinity for poly(C) repeats, more specifically, the affinity was higher for C-tetrads than for C-triplets. Protein modeling was performed based on the crystal structure of a KH domain bound to a TCCCT DNA sequence, pdb file, 3VKE, and by using the Yasara software (http://www.yasara.org )
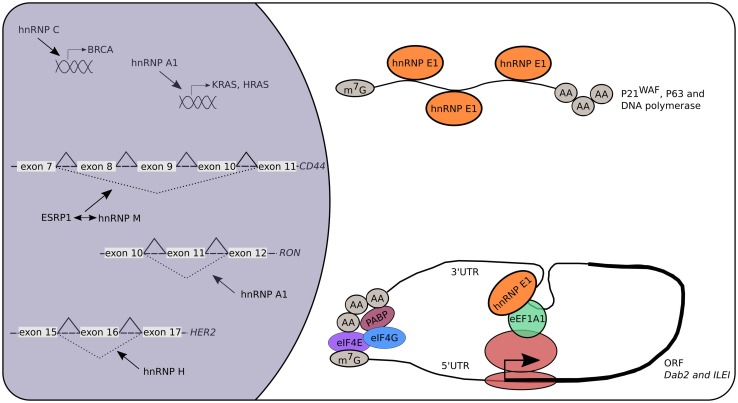
The role of hnRNPs in cancer. Multiple hnRNPs are linked to cancer and metastasis. Many of them act on the level of alternative splicing leading to truncated proteins because of exon skipping. This modulation can be direct (e.g., hnRNP A1 and H) or indirect (e.g., hnRNP M) by competing with other RBPs like ESRP1. Additionally, hnRNP A1 and C are known transcriptional regulators acting on the promotor of KRAS, HRAS and BRCA, respectively. Specific cytosolic functions of hnRNP E1 are linked to the initiation and progression of cancers. More specifically, hnRNP E1 controls the mRNA stability and therefore mRNA half-life of specific genes (p21WAF, p63, DNA polymerase η) linked to cancer development. Recently, the discovery that hnRNP E1 is a key player in the TGF-β-mediated EMT resulted in deciphering RBP components involved in the translational control of known oncogenes like Dab-2 and ILEI
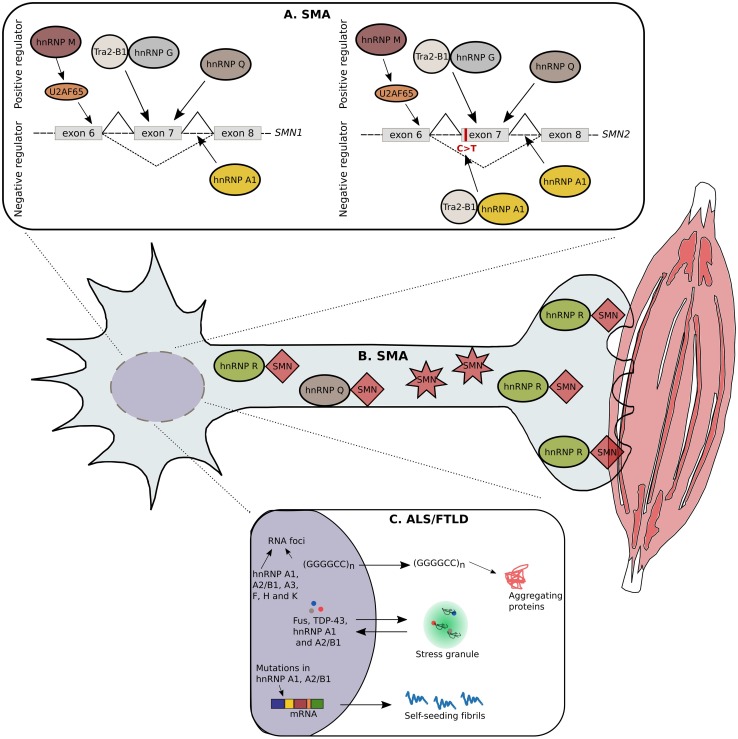
hnRNPs involved in SMA and ALS/FTLD. Due to extensive research in the last years, numerous hnRNPs are linked with SMA and ALS/FTLD. A single nucleotide substitution in the 5′-end of exon 7 in the SMN2 gene leads to exon skipping and consequently to a non-functional SMN protein. The hnRNP G can form a complex with the positive modifier, Tra2-β1, resulting in the retention of exon 7. In addition, hnRNP Q can bind to the single nucleotide substitution, thereby avoiding exon 7 skipping. The hnRNP M targets a splicing enhancer on exon 7 through the recruitment of U2AF65, leading to the production of a full-length transcript. In contrast, hnRNP A1 is found to be a negative regulator by binding to the splicing silencer, created by the single nucleotide substitution in SMN2. In addition, hnRNP A1 can bind to regions located in the introns of SMN1/2 (a). Besides being involved in alternative splicing, the hnRNP Q forms protein interactions with wild-type SMN, but is unable to bind to the truncated form. Similarly, the hnRNP R interacts with SMN in the cytosol of motor neurons; more specifically this interaction is present in the presynapses of neuromuscular junctions (b). C9orf72 repeat expansions causing ALS/FTLD leads to the formation of RNA foci containing hnRNP A1, A2/B1, A3, F, H and K suggesting RNA toxicity via the sequestration of RBPs as a possible pathomechanism. Furthermore, hnRNP A1 and A2/B1 are sequestered in cytosolic stress granules together with other RBPs causative for ALS/FTLD, such as FUS and TDP-43. Mutations found in the prion-like domain of hnRNP A1 and A2/B1 are causative for ALS/FTLD. In normal conditions, these hnRNPs have an intrinsic tendency to self-aggregate, because of their prion-like domains, but this tendency is abnormally increased because of the disease-causing mutations (c)
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
