

A voltage-dependent chloride channel fine-tunes photosynthesis in plants
- PMID: 27216227
- PMCID: PMC4890181
- DOI: 10.1038/ncomms11654
A voltage-dependent chloride channel fine-tunes photosynthesis in plants
Abstract
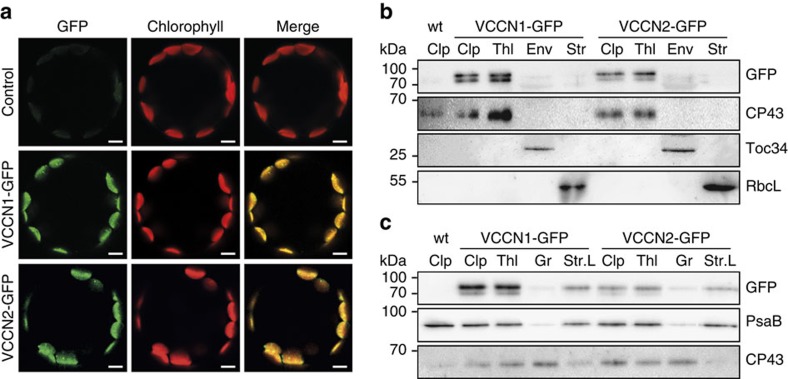
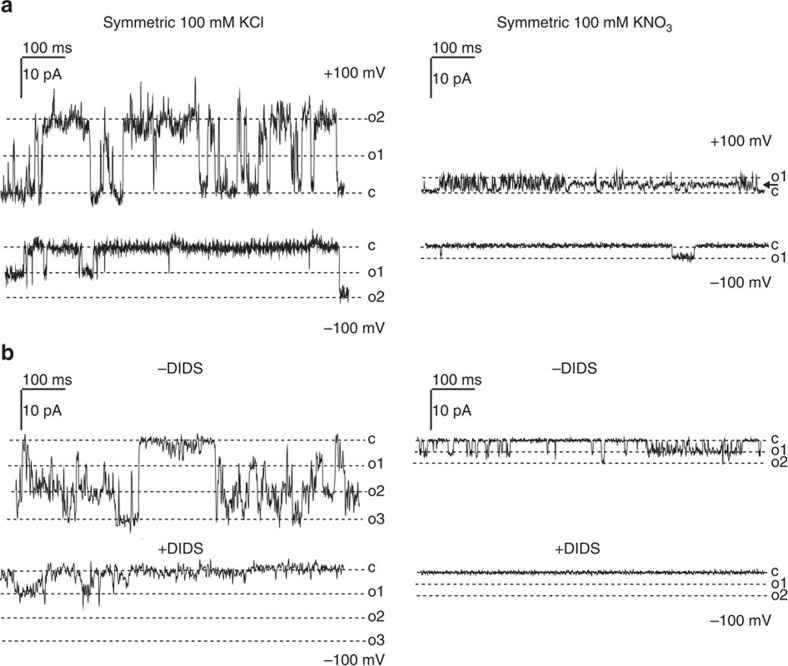
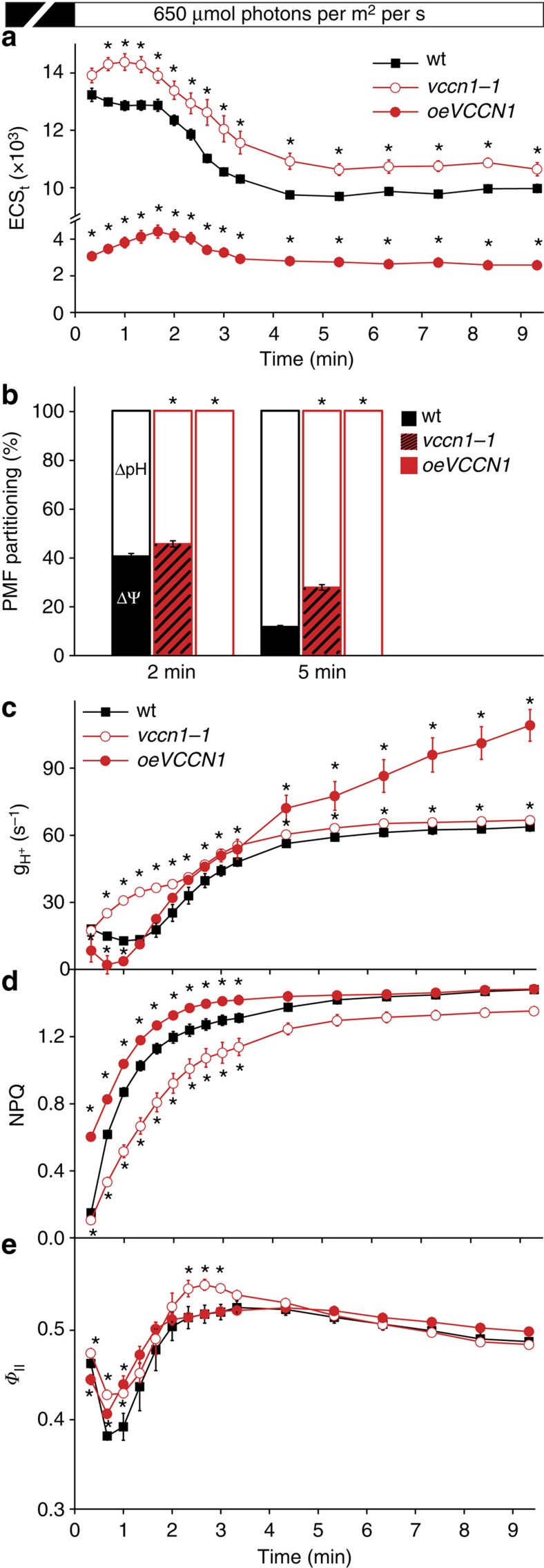
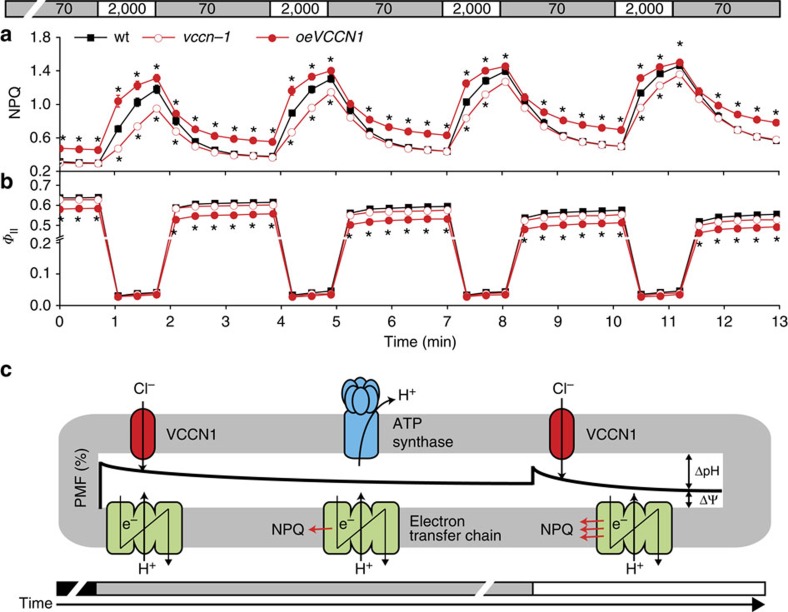
In natural habitats, plants frequently experience rapid changes in the intensity of sunlight. To cope with these changes and maximize growth, plants adjust photosynthetic light utilization in electron transport and photoprotective mechanisms. This involves a proton motive force (PMF) across the thylakoid membrane, postulated to be affected by unknown anion (Cl(-)) channels. Here we report that a bestrophin-like protein from Arabidopsis thaliana functions as a voltage-dependent Cl(-) channel in electrophysiological experiments. AtVCCN1 localizes to the thylakoid membrane, and fine-tunes PMF by anion influx into the lumen during illumination, adjusting electron transport and the photoprotective mechanisms. The activity of AtVCCN1 accelerates the activation of photoprotective mechanisms on sudden shifts to high light. Our results reveal that AtVCCN1, a member of a conserved anion channel family, acts as an early component in the rapid adjustment of photosynthesis in variable light environments.
Figures
References
-
- Finazzi G. et al.. Ions channels/transporters and chloroplast regulation. Cell Calcium 58, 86–97 (2014). - PubMed
-
- Pribil M., Labs M. & Leister D. Structure and dynamics of thylakoids in land plants. J. Exp. Bot. 65, 1955–1972 (2014). - PubMed
-
- Segalla A., Szabó I., Costantini P. & Giacometti G. M. Study of the effect of ion channel modulators on photosynthetic oxygen evolution. J. Chem. Inf. Model 45, 1691–1700 (2005). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
