

Estimating changing contexts in schizophrenia
- PMID: 27217338
- PMCID: PMC4939701
- DOI: 10.1093/brain/aww095
Estimating changing contexts in schizophrenia
Abstract
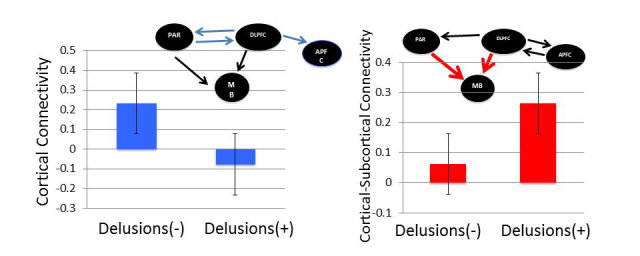
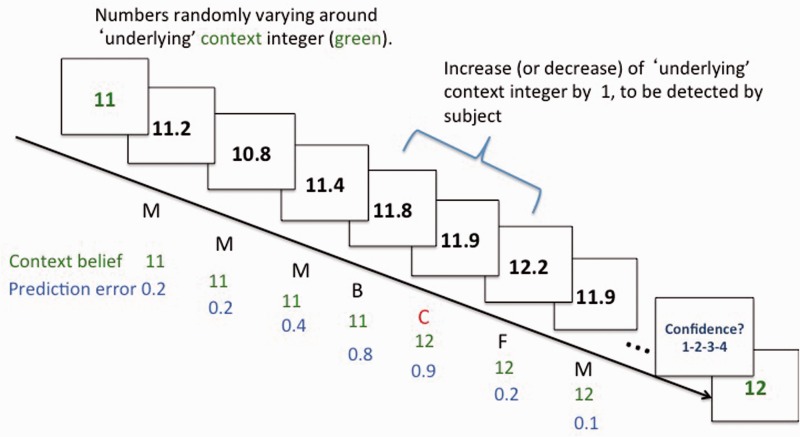
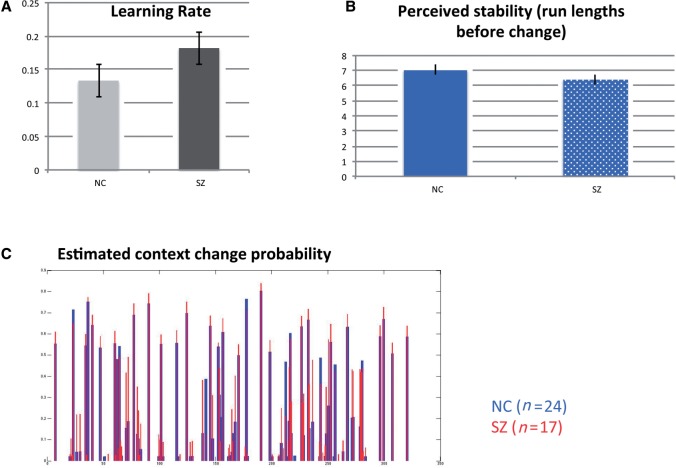
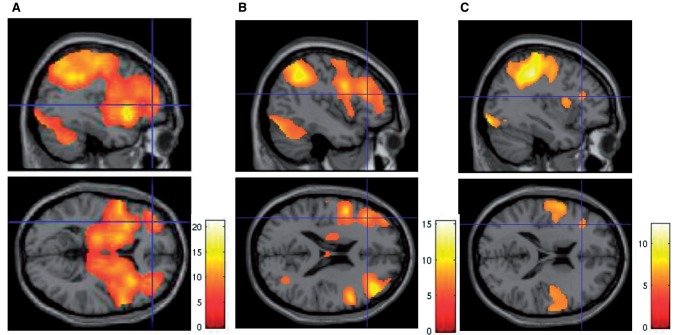
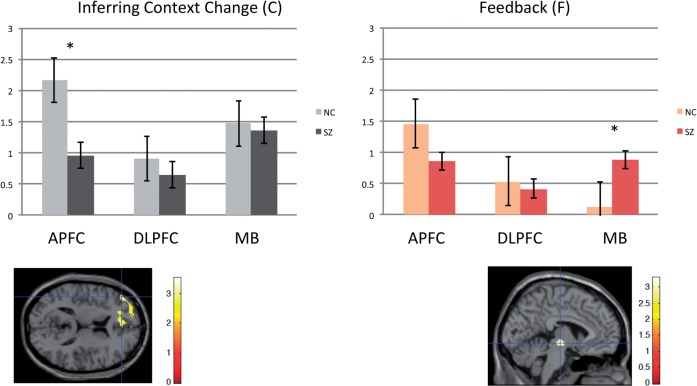
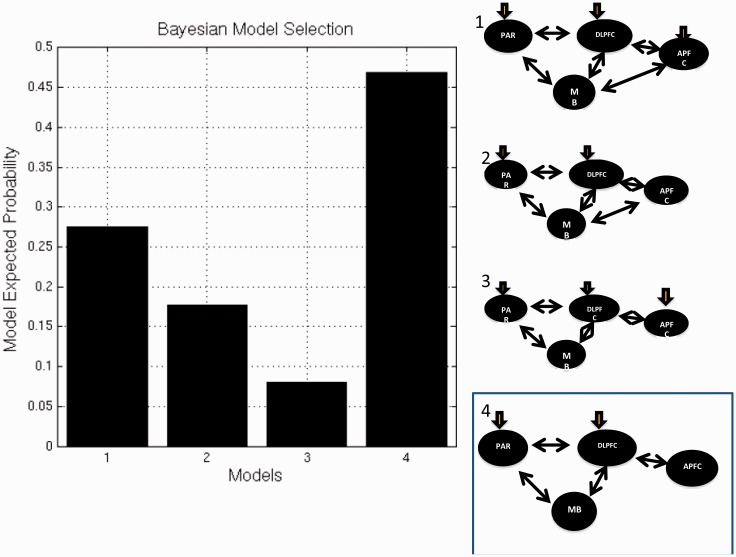
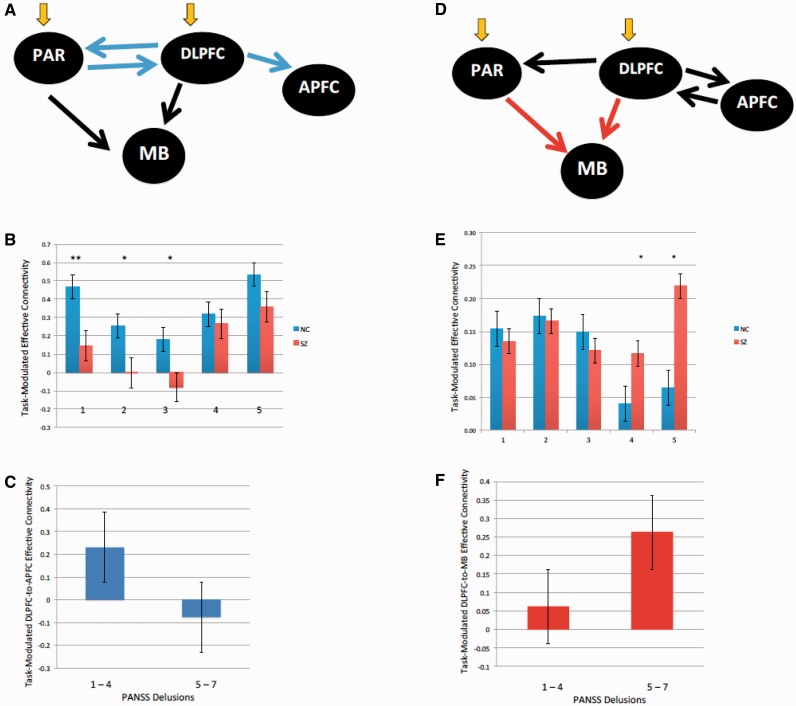
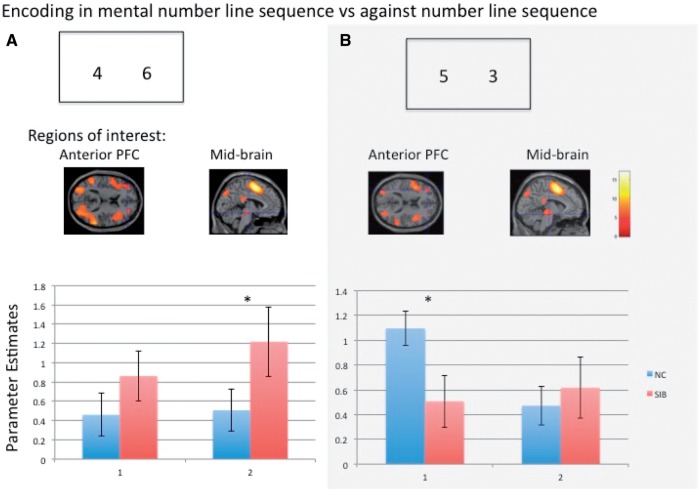
SEE STEPHAN ET AL DOI101093/AWW120 FOR A SCIENTIFIC COMMENTARY ON THIS WORK: Real world information is often abstract, dynamic and imprecise. Deciding if changes represent random fluctuations, or alterations in underlying contexts involve challenging probability estimations. Dysfunction may contribute to erroneous beliefs, such as delusions. Here we examined brain function during inferences about context change from noisy information. We examined cortical-subcortical circuitry engaging anterior and dorsolateral prefrontal cortex, and midbrain. We hypothesized that schizophrenia-related deficits in prefrontal function might overestimate context change probabilities, and that this more chaotic worldview may subsequently gain familiarity and be over-reinforced, with implications for delusions. We then examined these opposing information processing biases against less expected versus familiar information patterns in relation to genetic risk for schizophrenia in unaffected siblings. In one experiment, 17 patients with schizophrenia and 24 normal control subjects were presented in 3 T magnetic resonance imaging with numerical information varying noisily about a context integer, which occasionally shifted up or down. Subjects were to indicate when the inferred numerical context had changed. We fitted Bayesian models to estimate probabilities associated with change inferences. Dynamic causal models examined cortical-subcortical circuitry interactions at context change inference, and at subsequent reduced uncertainty. In a second experiment, genetic risk for schizophrenia associated with similar cortical-subcortical findings were explored in an independent sample of 36 normal control subjects and 35 unaffected siblings during processing of intuitive number sequences along the number line, or during the inverse, less familiar, sequence. In the first experiment, reduced Bayesian models fitting subject behaviour suggest that patients with schizophrenia overestimated context change probabilities. Here, patients engaged anterior prefrontal cortex relatively less than healthy controls, in part driven by reduced effective connectivity from dorsolateral prefrontal cortex to anterior prefrontal cortex. In processing subsequent information indicating reduced uncertainty of their predictions, patients engaged relatively increased mid-brain activation, driven in part by increased dorsolateral prefrontal cortex to midbrain connectivity. These dissociable reduced and exaggerated prefrontal and subcortical circuit functions were accentuated in patients with delusions. In the second experiment, analogous dissociable reduced anterior prefrontal cortex and exaggerated midbrain engagement occurred in unaffected siblings when processing less expected versus more familiar number sequences. In conclusion, patients overestimated ambiguous context change probabilities with relatively reduced anterior frontal engagement. Subsequent reduced uncertainty about contextual state appeared over-reinforced, potentially contributing to confirmation bias and a cascade of aberrant belief processing about a more chaotic world relevant to delusions. These opposing cortical-subcortical effects relate in part to genetic risk for schizophrenia, with analogous imbalances in neural processing of less expected versus familiar information patterns.
Keywords: anterior prefrontal cortex; context; delusions; effective connectivity; schizophrenia.
© The Author (2016). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
Comment in
-
Bayesian inference, dysconnectivity and neuromodulation in schizophrenia.Brain. 2016 Jul;139(Pt 7):1874-6. doi: 10.1093/brain/aww120. Brain. 2016. PMID: 27343221 No abstract available.
References
-
- Balzan R, Delfabbro P, Galletly C, Woodward T. Confirmation biases across the psychosis continuum: the contribution of hypersalient evidence-hypothesis matches. Br J Clin Psychol 2013; 52: 53–69. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
