

Polyhydroxyalkanoate (PHA) Granules Have no Phospholipids
- PMID: 27222167
- PMCID: PMC4879537
- DOI: 10.1038/srep26612
Polyhydroxyalkanoate (PHA) Granules Have no Phospholipids
Abstract
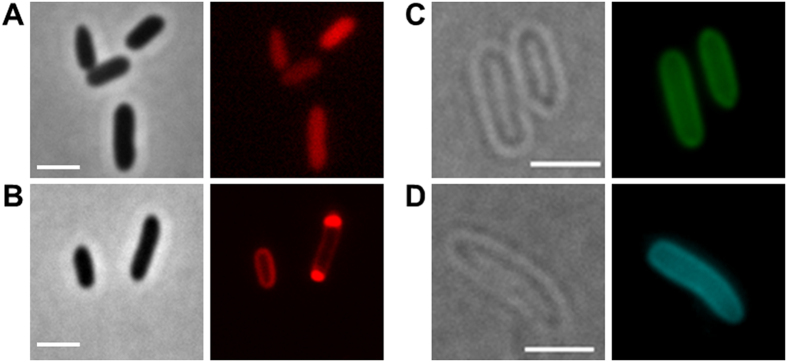
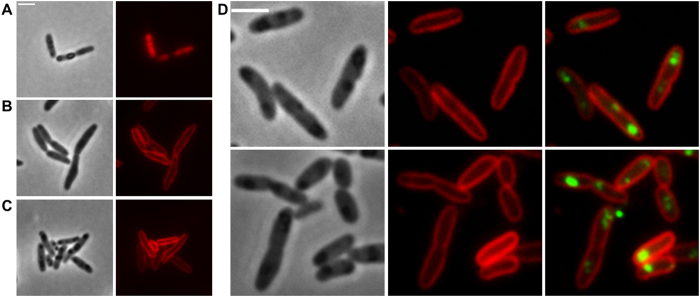
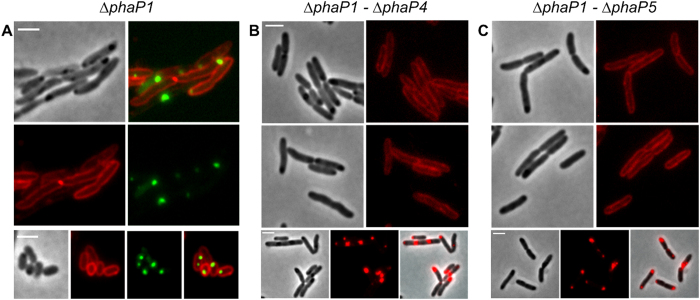
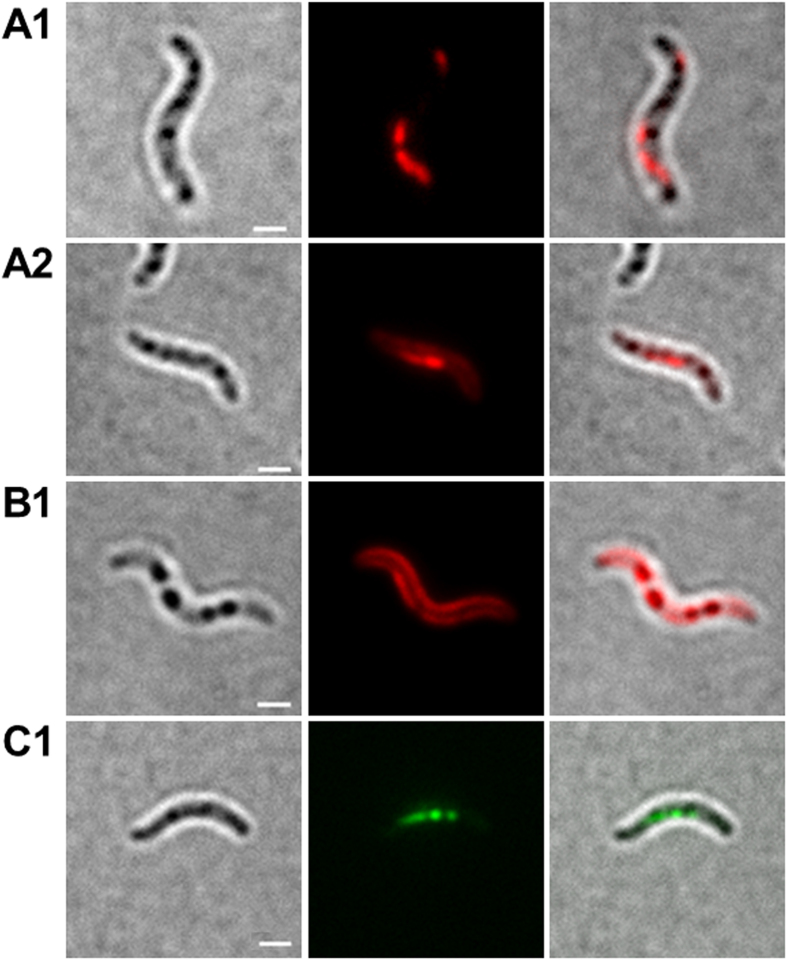
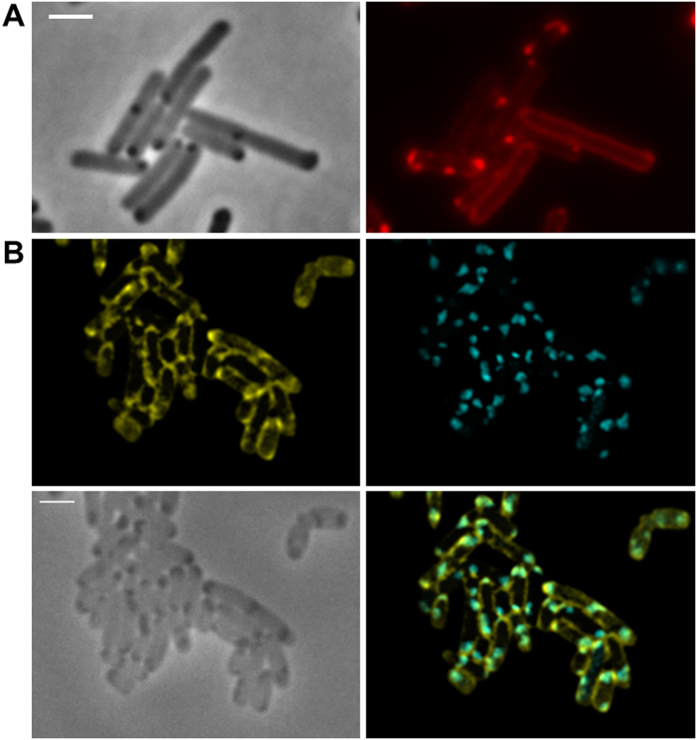
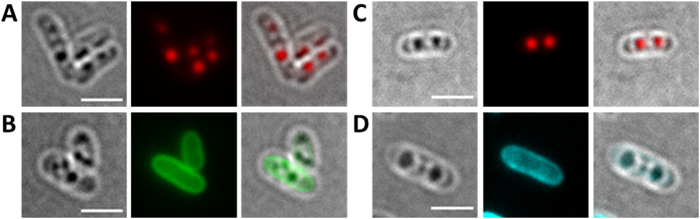
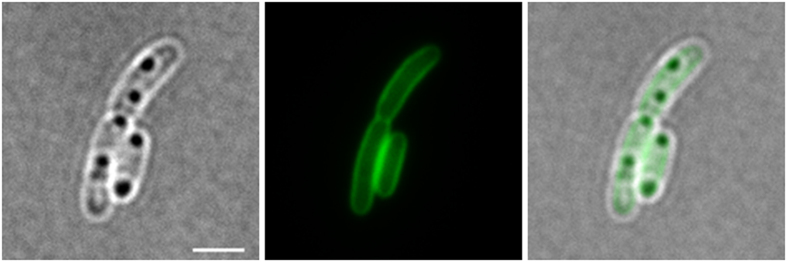
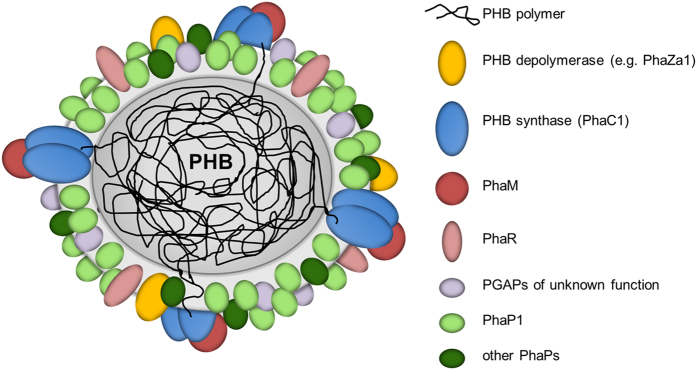
Polyhydroxybutyrate (PHB) granules, also designated as carbonosomes, are supra-molecular complexes in prokaryotes consisting of a PHB polymer core and a surface layer of structural and functional proteins. The presence of suspected phospholipids in the surface layer is based on in vitro data of isolated PHB granules and is often shown in cartoons of the PHB granule structure in reviews on PHB metabolism. However, the in vivo presence of a phospholipid layer has never been demonstrated. We addressed this topic by the expression of fusion proteins of DsRed2EC and other fluorescent proteins with the phospholipid-binding domain (LactC2) of lactadherin in three model organisms. The fusion proteins specifically localized at the cell membrane of Ralstonia eutropha but did not co-localize with PHB granules. The same result was obtained for Pseudomonas putida, a species that accumulates another type of polyhydroxyalkanoate (PHA) granules related to PHB. Notably, DsRed2EC-LactC2 expressed in Magnetospirillum gryphiswaldense was detected at the position of membrane-enclosed magnetosome chains and at the cytoplasmic membrane but not at PHB granules. In conclusion, the carbonosomes of representatives of α-proteobacteria, β-proteobacteria and γ-proteobacteria have no phospholipids in vivo and we postulate that the PHB/PHA granule surface layers in natural producers generally are free of phospholipids and consist of proteins only.
Figures
References
-
- Stubbe J. et al. Nontemplate-dependent polymerization processes: polyhydroxyalkanoate synthases as a paradigm. Annu. Rev. Biochem. 74, 433–480 (2005). - PubMed
-
- Pötter M. & Steinbüchel A. Biogenesis and Structure of polyhydroxyalkanoate granules. Microbiol. Monogr. 1, 110–136 (2006).
-
- Grage K. et al. Bacterial polyhydroxyalkanoate granules: biogenesis, structure, and potential use as nano-/micro-beads in biotechnological and biomedical applications. Biomacromolecules 10, 660–669 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
