

NPR-9, a Galanin-Like G-Protein Coupled Receptor, and GLR-1 Regulate Interneuronal Circuitry Underlying Multisensory Integration of Environmental Cues in Caenorhabditis elegans
- PMID: 27223098
- PMCID: PMC4880332
- DOI: 10.1371/journal.pgen.1006050
NPR-9, a Galanin-Like G-Protein Coupled Receptor, and GLR-1 Regulate Interneuronal Circuitry Underlying Multisensory Integration of Environmental Cues in Caenorhabditis elegans
Abstract
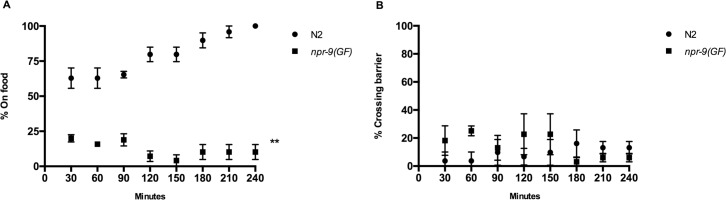
C. elegans inhabit environments that require detection of diverse stimuli to modulate locomotion in order to avoid unfavourable conditions. In a mammalian context, a failure to appropriately integrate environmental signals can lead to Parkinson's, Alzheimer's, and epilepsy. Provided that the circuitry underlying mammalian sensory integration can be prohibitively complex, we analyzed nematode behavioral responses in differing environmental contexts to evaluate the regulation of context dependent circuit reconfiguration and sensorimotor control. Our work has added to the complexity of a known parallel circuit, mediated by interneurons AVA and AIB, that integrates sensory cues and is responsible for the initiation of backwards locomotion. Our analysis of the galanin-like G-protein coupled receptor NPR-9 in C. elegans revealed that upregulation of galanin signaling impedes the integration of sensory evoked neuronal signals. Although the expression pattern of npr-9 is limited to AIB, upregulation of the receptor appears to impede AIB and AVA circuits to broadly prevent backwards locomotion, i.e. reversals, suggesting that these two pathways functionally interact. Galanin signaling similarly plays a broadly inhibitory role in mammalian models. Moreover, our identification of a mutant, which rarely initiates backwards movement, allowed us to interrogate locomotory mechanisms underlying chemotaxis. In support of the pirouette model of chemotaxis, organisms that did not exhibit reversal behavior were unable to navigate towards an attractant peak. We also assessed ionotropic glutamate receptor GLR-1 cell-specifically within AIB and determined that GLR-1 fine-tunes AIB activity to modify locomotion following reversal events. Our research highlights that signal integration underlying the initiation and fine-tuning of backwards locomotion is AIB and NPR-9 dependent, and has demonstrated the suitability of C. elegans for analysis of multisensory integration and sensorimotor control.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
-
- White JG, Southgate E, Thomson JN, Brenner S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos Trans R Soc Lond B Biol Sci 1986. November 12;314(1165):1–340. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
