

Expression of transmembrane protein 26 (TMEM26) in breast cancer and its association with drug response
- PMID: 27224909
- PMCID: PMC5122400
- DOI: 10.18632/oncotarget.9493
Expression of transmembrane protein 26 (TMEM26) in breast cancer and its association with drug response
Abstract
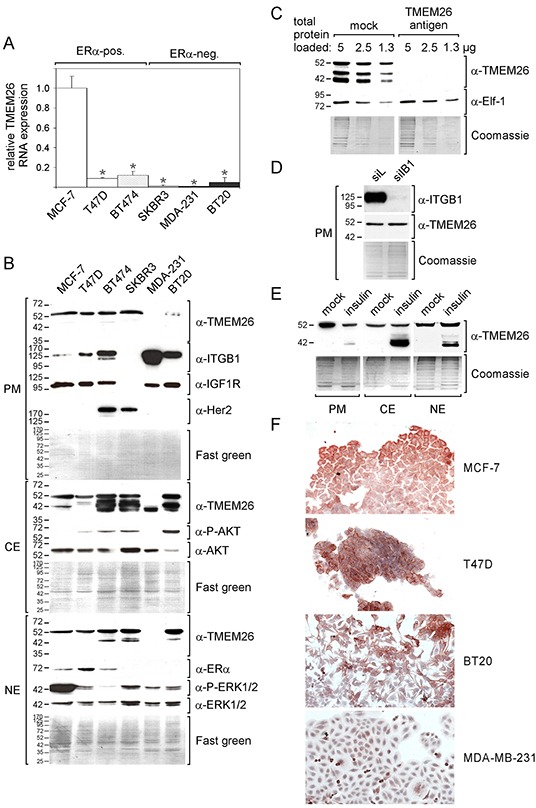
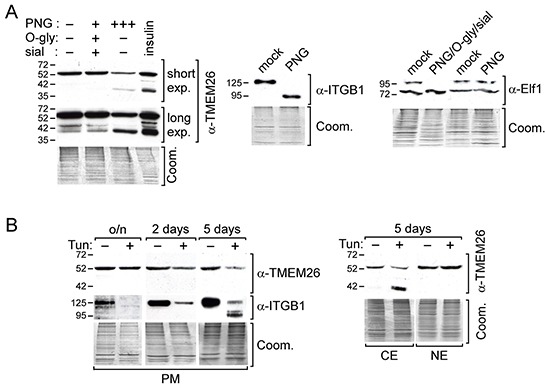
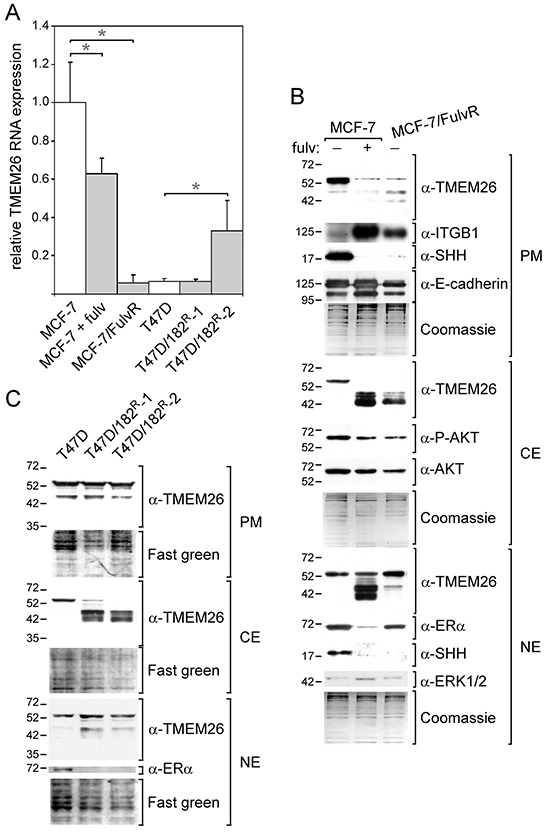
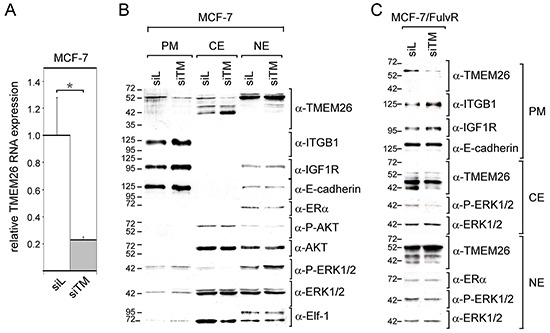
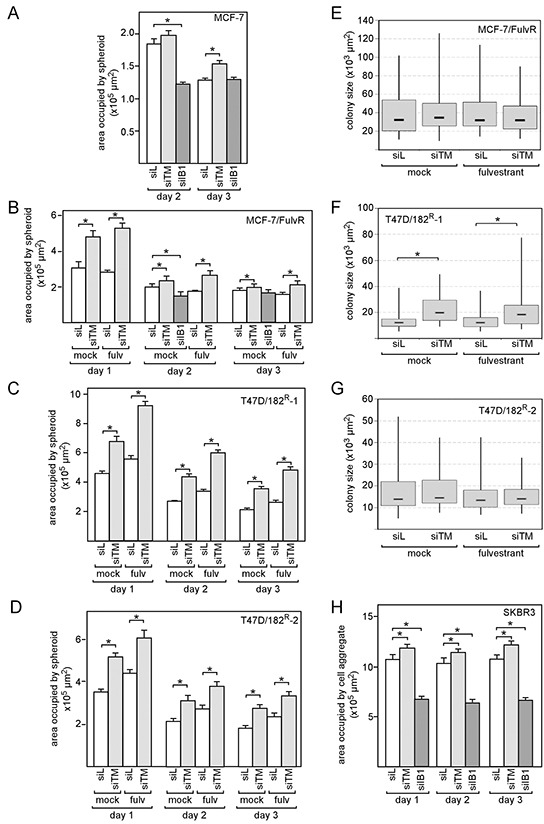
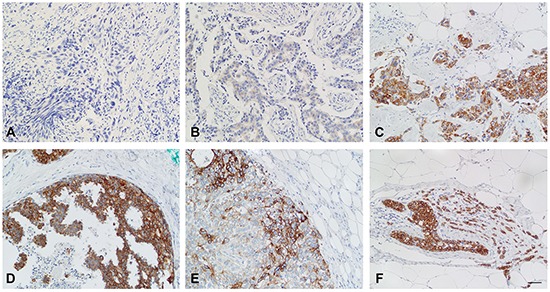
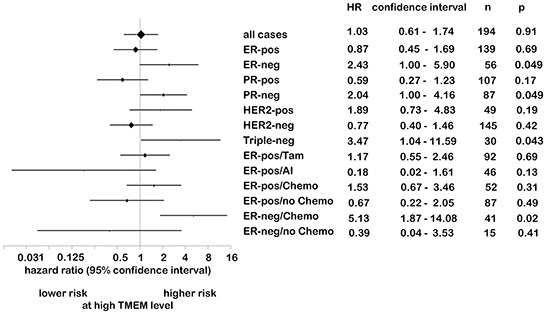
We have previously shown that stromal cells desensitize breast cancer cells to the anti-estrogen fulvestrant and, along with it, downregulate the expression of TMEM26 (transmembrane protein 26). In an effort to study the function and regulation of TMEM26 in breast cancer cells, we found that breast cancer cells express non-glycosylated and N-glycosylated isoforms of the TMEM26 protein and demonstrate that N-glycosylation is important for its retention at the plasma membrane. Fulvestrant induced significant changes in expression and in the N-glycosylation status of TMEM26. In primary breast cancer, TMEM26 protein expression was higher in ERα (estrogen receptor α)/PR (progesterone receptor)-positive cancers. These data suggest that ERα is a major regulator of TMEM26. Significant changes in TMEM26 expression and N-glycosylation were also found, when MCF-7 and T47D cells acquired fulvestrant resistance. Furthermore, patients who received aromatase inhibitor treatment tend to have a higher risk of recurrence when tumoral TMEM26 protein expression is low. In addition, TMEM26 negatively regulates the expression of integrin β1, an important factor involved in endocrine resistance. Data obtained by spheroid formation assays confirmed that TMEM26 and integrin β1 can have opposite effects in breast cancer cells. These data are consistent with the hypothesis that, in ERα-positive breast cancer, TMEM26 may function as a tumor suppressor by impeding the acquisition of endocrine resistance. In contrast, in ERα-negative breast cancer, particularly triple-negative cancer, high TMEM26 expression was found to be associated with a higher risk of recurrence. This implies that TMEM26 has different functions in ERα-positive and -negative breast cancer.
Keywords: anti-estrogen resistance; estrogen receptor; integrin beta1; sonic hedgehog; triple-negative breast cancer.
Conflict of interest statement
The authors declare there are no conflicts of interests.
Figures
Similar articles
-
Different epigenetic mechanisms of ERα implicated in the fate of fulvestrant-resistant breast cancer.J Steroid Biochem Mol Biol. 2017 Mar;167:115-125. doi: 10.1016/j.jsbmb.2016.11.017. Epub 2016 Nov 22. J Steroid Biochem Mol Biol. 2017. PMID: 27888136
-
Reprogramming of the ERRα and ERα target gene landscape triggers tamoxifen resistance in breast cancer.Cancer Res. 2015 Feb 15;75(4):720-31. doi: 10.1158/0008-5472.CAN-14-0652. Epub 2015 Feb 2. Cancer Res. 2015. PMID: 25643697
-
Stromal cells promote anti-estrogen resistance of breast cancer cells through an insulin-like growth factor binding protein 5 (IGFBP5)/B-cell leukemia/lymphoma 3 (Bcl-3) axis.Oncotarget. 2015 Nov 17;6(36):39307-28. doi: 10.18632/oncotarget.5624. Oncotarget. 2015. PMID: 26515727 Free PMC article.
-
Long Non-Coding RNA H19 Acts as an Estrogen Receptor Modulator that is Required for Endocrine Therapy Resistance in ER+ Breast Cancer Cells.Cell Physiol Biochem. 2018;51(4):1518-1532. doi: 10.1159/000495643. Epub 2018 Nov 29. Cell Physiol Biochem. 2018. PMID: 30497079
-
Rewiring of the Endocrine Network in Triple-Negative Breast Cancer.Front Oncol. 2022 Jun 30;12:830894. doi: 10.3389/fonc.2022.830894. eCollection 2022. Front Oncol. 2022. PMID: 35847875 Free PMC article. Review.
Cited by
-
The tamoxifen-regulated, long non-coding RNA LINC00992 affects proliferation, migration, and expression of tamoxifen resistance-associated genes in MCF-7 breast cancer cells.Contemp Oncol (Pozn). 2022;26(4):294-305. doi: 10.5114/wo.2023.125000. Epub 2022 Dec 30. Contemp Oncol (Pozn). 2022. PMID: 36816389 Free PMC article.
-
Identification and Validation of Immune-Related Long Non-Coding RNA Signature for Predicting Immunotherapeutic Response and Prognosis in NSCLC Patients Treated With Immunotherapy.Front Oncol. 2022 Jul 4;12:899925. doi: 10.3389/fonc.2022.899925. eCollection 2022. Front Oncol. 2022. PMID: 35860577 Free PMC article.
-
Combining Pathway Identification and Breast Cancer Survival Prediction via Screening-Network Methods.Front Genet. 2018 Jun 14;9:206. doi: 10.3389/fgene.2018.00206. eCollection 2018. Front Genet. 2018. PMID: 29963073 Free PMC article.
-
Genome-Wide Association Study of H/L Traits in Chicken.Animals (Basel). 2019 May 21;9(5):260. doi: 10.3390/ani9050260. Animals (Basel). 2019. PMID: 31117270 Free PMC article.
-
Long-term exposure to carcinoma-associated fibroblasts makes breast cancer cells addictive to integrin β1.Oncotarget. 2018 Apr 24;9(31):22079-22094. doi: 10.18632/oncotarget.25183. eCollection 2018 Apr 24. Oncotarget. 2018. PMID: 29774124 Free PMC article.
References
-
- Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87–108. - PubMed
-
- Davies C, Godwin J, Gray R, Clarke M, Cutter D, Darby S, McGale P, Pan HC, Taylor C, Wang YC, Dowsett M, Ingle J, Peto R. Relevance of breast cancer hormone receptors and other factors to the efficacy of adjuvant tamoxifen: patient-level meta-analysis of randomised trials. Lancet. 2011;378:771–784. - PMC - PubMed
-
- Hudis CA. Trastuzumab - mechanism of action and use in clinical practice. N Engl J Med. 2007;357:39–51. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
