

TIM-4 is differentially expressed in the distinct subsets of dendritic cells in skin and skin-draining lymph nodes and controls skin Langerhans cell homeostasis
- PMID: 27224924
- PMCID: PMC5122327
- DOI: 10.18632/oncotarget.9546
TIM-4 is differentially expressed in the distinct subsets of dendritic cells in skin and skin-draining lymph nodes and controls skin Langerhans cell homeostasis
Abstract
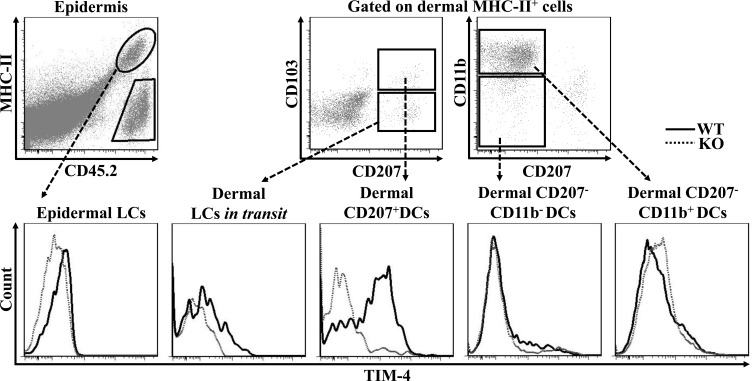
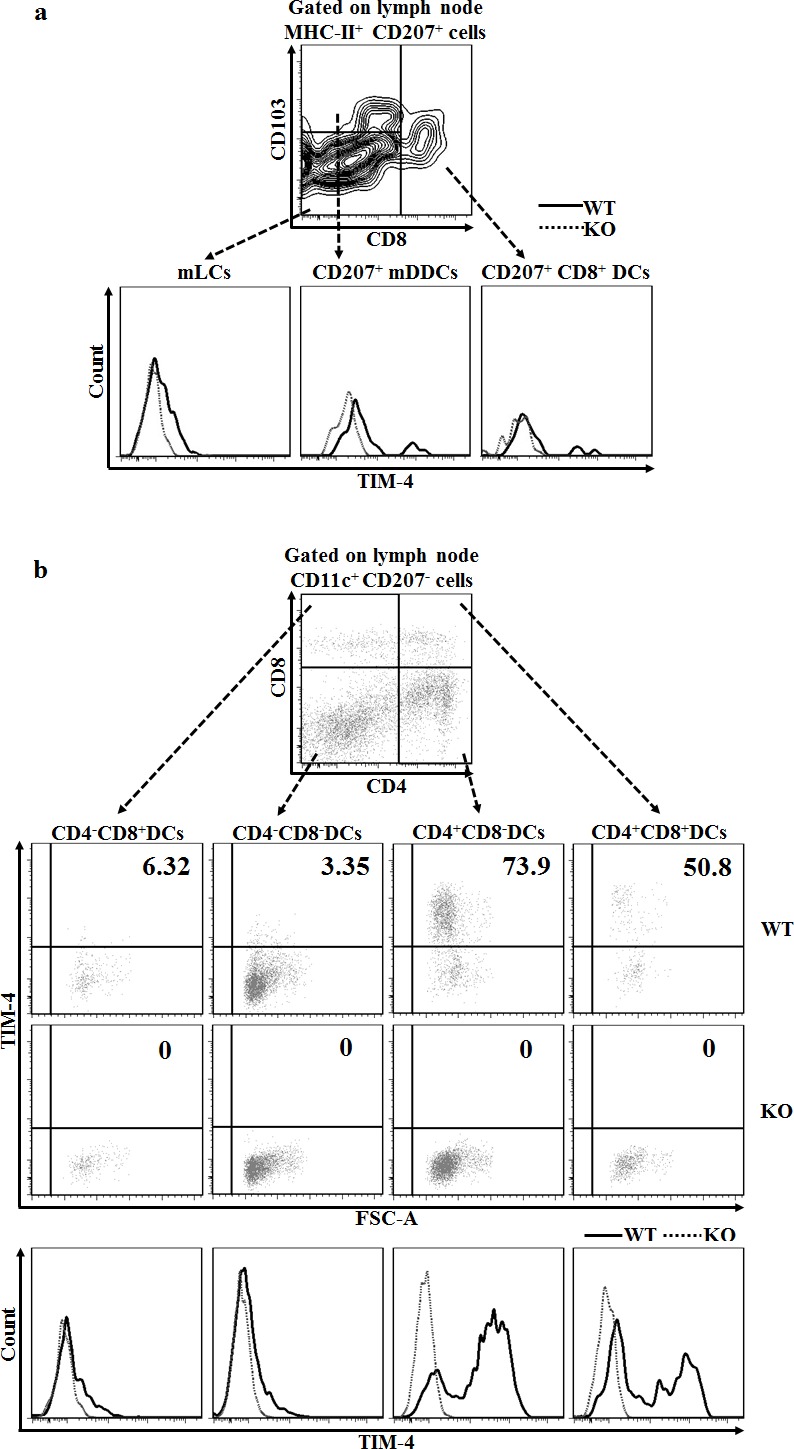
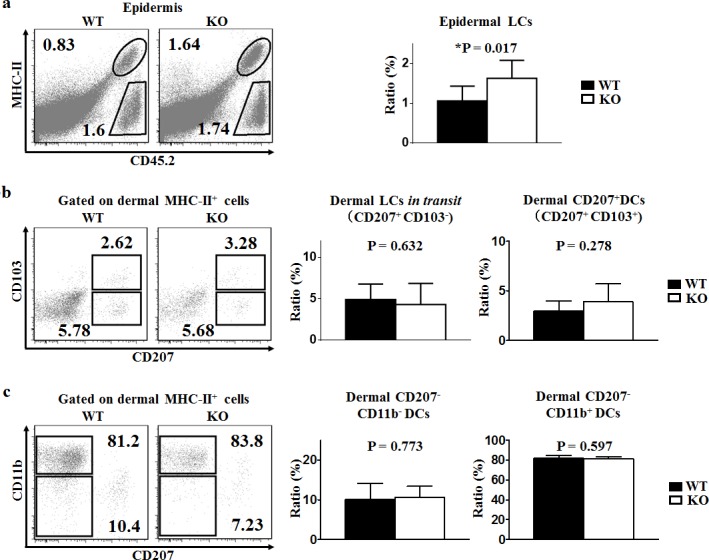
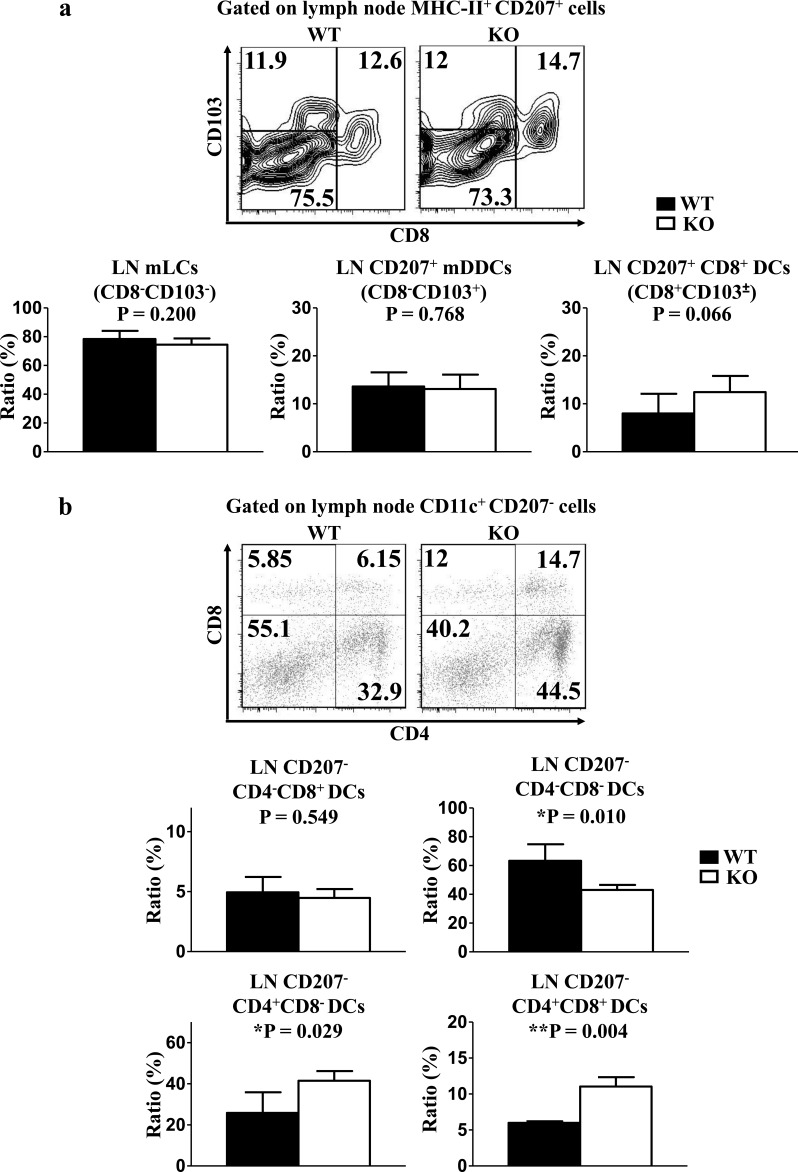
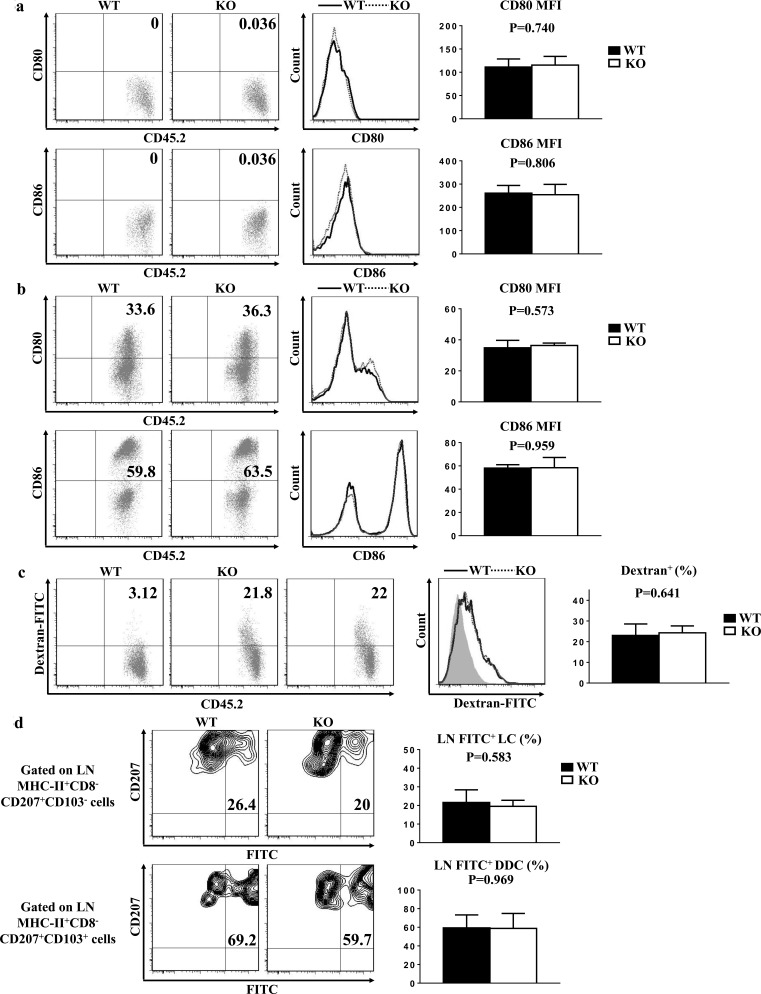
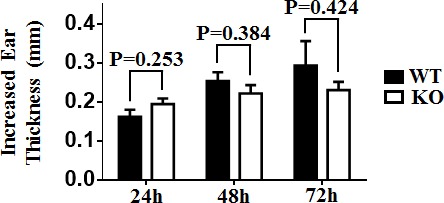
T cell immunoglobulin and mucin-4 (TIM-4), mainly expressed on dendritic cells (DC) and macrophages, plays an essential role in regulating immune responses. Langerhans cells (LC), which are the sole DC subpopulation residing at the epidermis, are potent mediators of immune surveillance and tolerance. However, the significance of TIM-4 on epidermal LCs, along with other cutaneous DCs, remains totally unexplored. For the first time, we discovered that epidermal LCs expressed TIM-4 and displayed an increased level of TIM-4 expression upon migration. We also found that dermal CD207+ DCs and lymph node (LN) resident CD207-CD4+ DCs highly expressed TIM-4, while dermal CD207- DCs and LN CD207-CD4- DCs had limited TIM-4 expressions. Using TIM-4-deficient mice, we further demonstrated that loss of TIM-4 significantly upregulated the frequencies of epidermal LCs and LN resident CD207-CD4+ DCs. In spite of this, the epidermal LCs of TIM-4-deficient mice displayed normal phagocytic and migratory abilities, comparable maturation status upon the stimulation as well as normal repopulation under the inflamed state. Moreover, lack of TIM-4 did not affect dinitrofluorobenzene-induced contact hypersensitivity response. In conclusion, our results indicated that TIM-4 was differentially expressed in the distinct subsets of DCs in skin and skin-draining LNs, and specifically regulated epidermal LC and LN CD207-CD4+ DC homeostasis.
Keywords: Immune response; Immunity; Immunology and Microbiology Section; Langerhans cells; TIM-4; dendritic cells; skin; skin-draining lymph node.
Conflict of interest statement
The authors state no conflict of interest.
Figures

Similar articles
-
Dynamics and function of Langerhans cells in vivo: dermal dendritic cells colonize lymph node areas distinct from slower migrating Langerhans cells.Immunity. 2005 May;22(5):643-54. doi: 10.1016/j.immuni.2005.04.004. Immunity. 2005. PMID: 15894281
-
Fate mapping analysis reveals a novel murine dermal migratory Langerhans-like cell population.Elife. 2021 Mar 26;10:e65412. doi: 10.7554/eLife.65412. Elife. 2021. PMID: 33769279 Free PMC article.
-
Osteopontin is involved in the initiation of cutaneous contact hypersensitivity by inducing Langerhans and dendritic cell migration to lymph nodes.J Exp Med. 2001 Nov 5;194(9):1219-29. doi: 10.1084/jem.194.9.1219. J Exp Med. 2001. PMID: 11696588 Free PMC article.
-
Langerhans cells--revisiting the paradigm using genetically engineered mice.Trends Immunol. 2006 Mar;27(3):132-9. doi: 10.1016/j.it.2006.01.003. Epub 2006 Feb 3. Trends Immunol. 2006. PMID: 16458606 Review.
-
Dendritic cells and T cells in the regulation of cutaneous immunity.Adv Dermatol. 2007;23:307-33. doi: 10.1016/j.yadr.2007.07.014. Adv Dermatol. 2007. PMID: 18159907 Review.
Cited by
-
Noncanonical autophagy in dermal dendritic cells mediates immunosuppressive effects of UV exposure.J Allergy Clin Immunol. 2020 May;145(5):1389-1405. doi: 10.1016/j.jaci.2019.11.041. Epub 2019 Dec 16. J Allergy Clin Immunol. 2020. Retraction in: J Allergy Clin Immunol. 2022 Oct;150(4):979. doi: 10.1016/j.jaci.2022.08.006. PMID: 31837371 Free PMC article. Retracted.
-
Having an Old Friend for Dinner: The Interplay between Apoptotic Cells and Efferocytes.Cells. 2021 May 20;10(5):1265. doi: 10.3390/cells10051265. Cells. 2021. PMID: 34065321 Free PMC article. Review.
-
Single-cell RNA-Seq reveals the transcriptional landscape and heterogeneity of skin macrophages in Vsir-/- murine psoriasis.Theranostics. 2020 Aug 21;10(23):10483-10497. doi: 10.7150/thno.45614. eCollection 2020. Theranostics. 2020. PMID: 32929361 Free PMC article.
-
IL-4/IL-13 polarization of macrophages enhances Ebola virus glycoprotein-dependent infection.PLoS Negl Trop Dis. 2019 Dec 11;13(12):e0007819. doi: 10.1371/journal.pntd.0007819. eCollection 2019 Dec. PLoS Negl Trop Dis. 2019. PMID: 31825972 Free PMC article.
-
Ameliorative effects of sea buckthorn oil on DNCB induced atopic dermatitis model mice via regulation the balance of Th1/Th2.BMC Complement Med Ther. 2020 Aug 26;20(1):263. doi: 10.1186/s12906-020-02997-2. BMC Complement Med Ther. 2020. PMID: 32843010 Free PMC article.
References
-
- McIntire JJ, Umetsu SE, Akbari O, Potter M, Kuchroo VK, Barsh GS, Freeman GJ, Umetsu DT, DeKruyff RH. Identification of Tapr (an airway hyperreactivity regulatory locus) and the linked Tim gene family. Nature immunology. 2001;2:1109–1116. - PubMed
-
- Kuchroo VK, Umetsu DT, DeKruyff RH, Freeman GJ. The TIM gene family: emerging roles in immunity and disease. Nature reviews Immunology. 2003;3:454–462. - PubMed
-
- Li Z, Ju Z, Frieri M. The T-cell immunoglobulin and mucin domain (Tim) gene family in asthma, allergy, and autoimmunity. Allergy and asthma proceedings. 2013;34:e21–26. - PubMed
-
- Rennert PD. Novel roles for TIM-1 in immunity and infection. Immunology letters. 2011;141:28–35. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
