

Deficiency of myeloid-related proteins 8 and 14 (Mrp8/Mrp14) does not block inflammaging but prevents steatosis
- PMID: 27224926
- PMCID: PMC5094943
- DOI: 10.18632/oncotarget.9550
Deficiency of myeloid-related proteins 8 and 14 (Mrp8/Mrp14) does not block inflammaging but prevents steatosis
Abstract
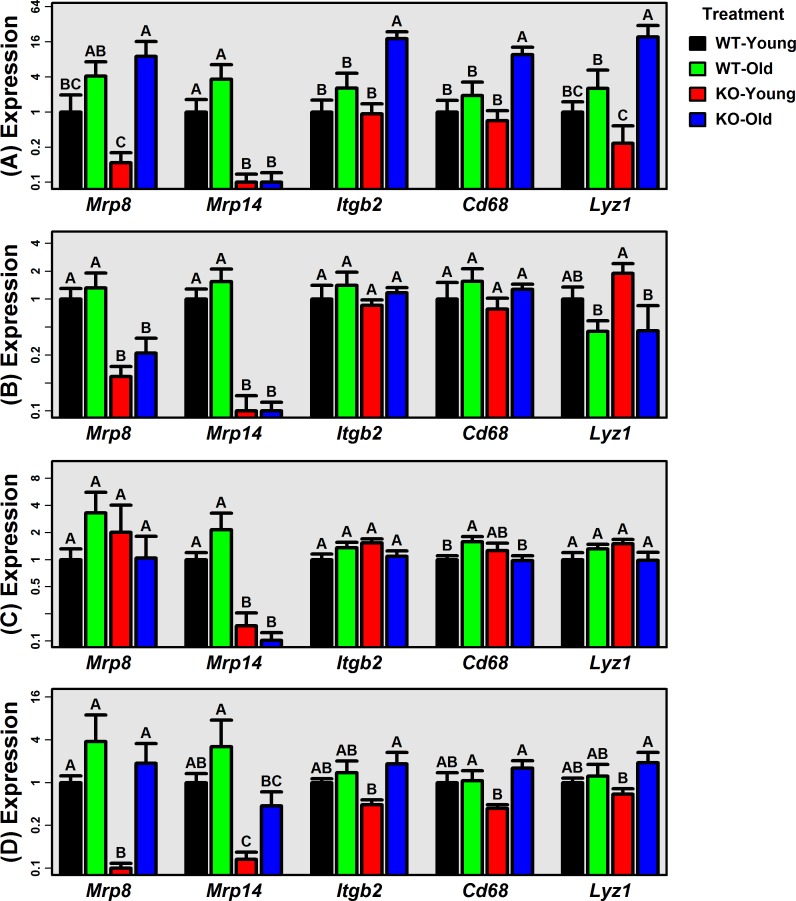
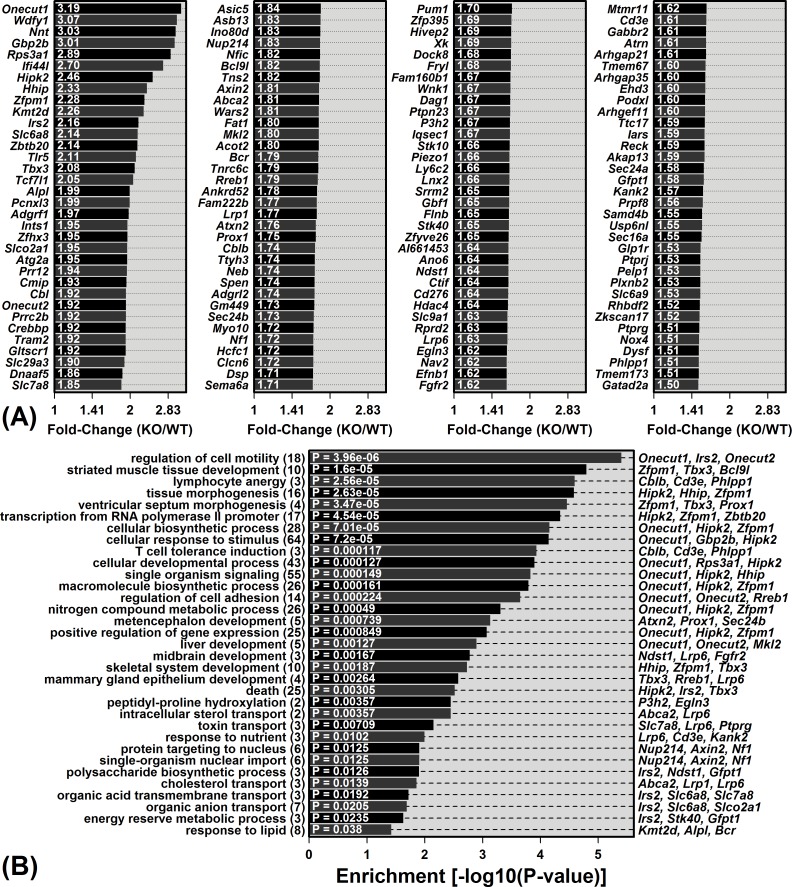
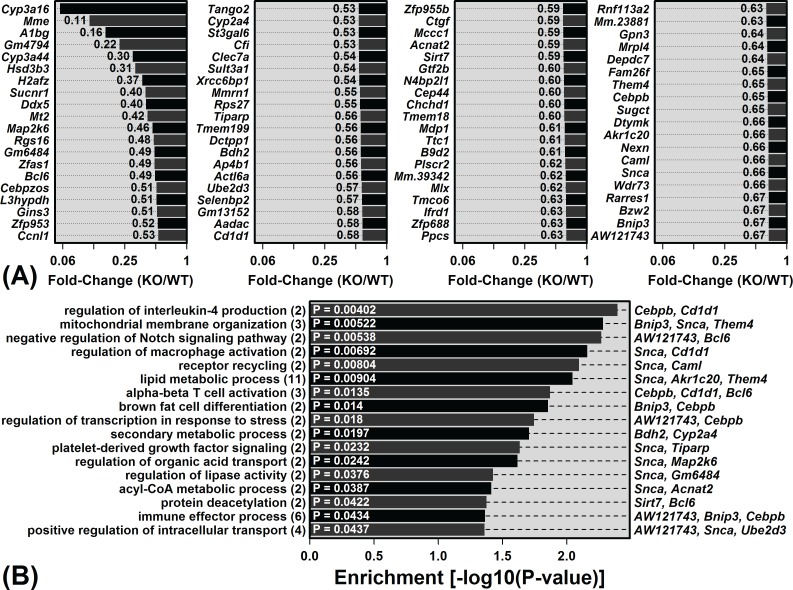
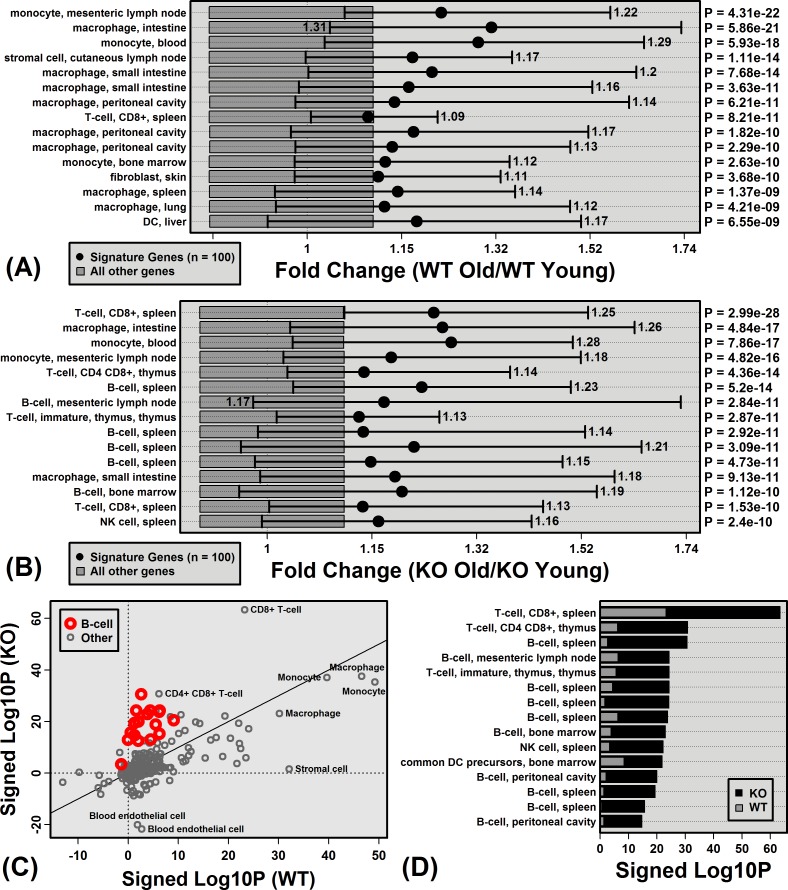
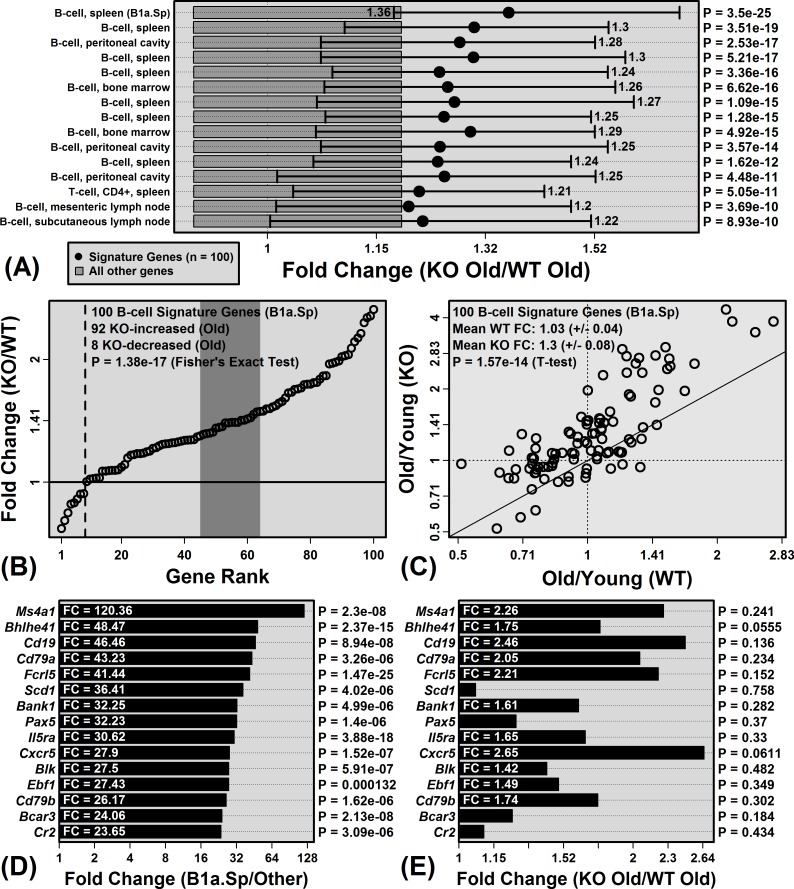
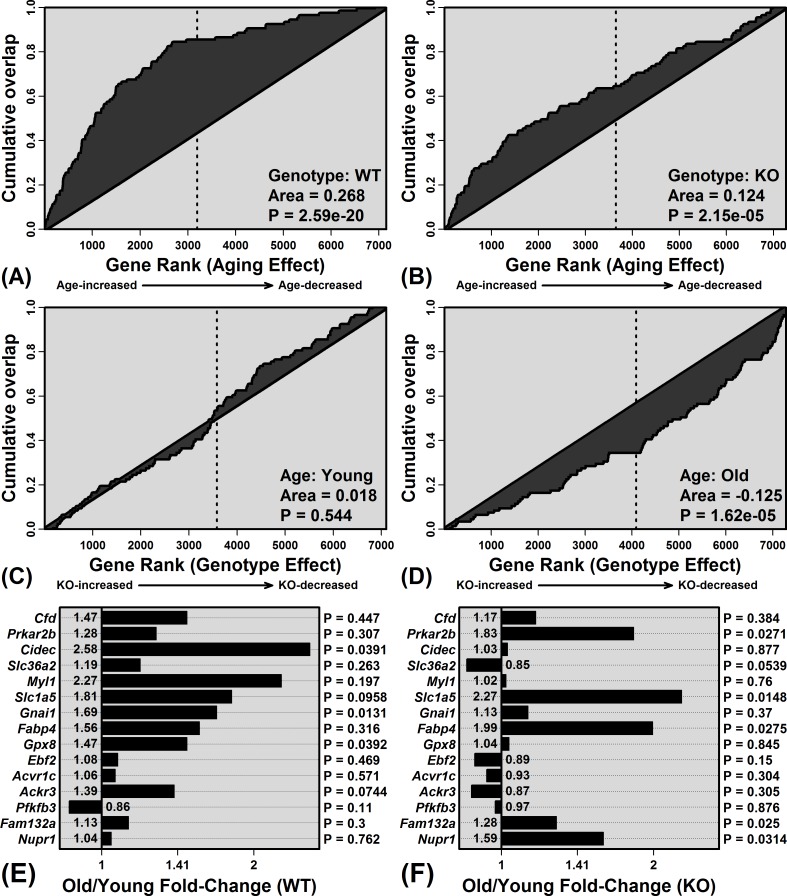
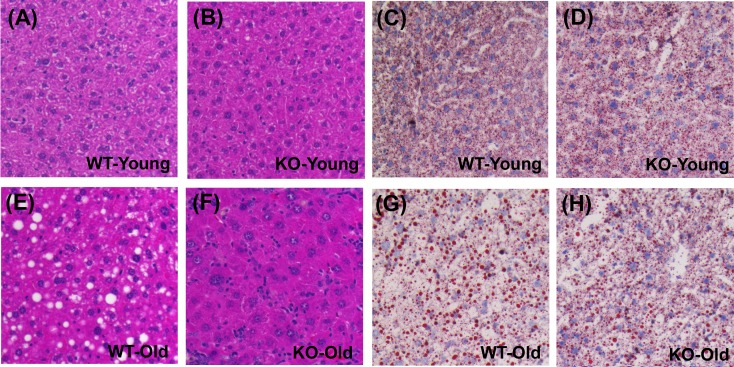
The Mrp8 and Mrp14 proteins (calprotectin) accumulate within tissues during aging and may contribute to chronic inflammation. To address this possibility, we evaluated female calprotectin-deficient Mrp14-KO and wild-type (WT) mice at 5 and 24 months of age. However, there was no evidence that age-related inflammation is blunted in KO mice. Inflammation markers were in fact elevated in livers from old KO mice, and microarray analysis revealed more consistent elevation of genes specifically expressed by B-cells and T-cells. Adipose-specific genes, however, were less consistently elevated in aged KO mice, suggesting an anti-steatosis effect of Mrp8/14 deficiency. Consistent with this, genes decreased by the anti-steatosis agent SRT1720 were decreased in old KO compared to old WT mice. Expression of lipid metabolism genes was altered in KO mice at 5 months of age, along with genes associated with development, biosynthesis and immunity. These early-age effects of Mrp8/14 deficiency, in the absence of any external stressor, were unexpected. Taken together, our findings demonstrate a pro-steatosis rather than pro-inflammatory role of calprotectin within the aging liver. This appears to reflect a developmental-metabolic phenotype of Mrp14-KO mice that is manifest at a young age in the absence of pro-inflammatory stimuli.
Keywords: B-cell; Gerotarget; S100a8; calgranulin; calprotectin; microarray.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
MRP14 is dispensable for LPS-induced shock in BALB/c mice.Immunol Lett. 2018 Feb;194:13-20. doi: 10.1016/j.imlet.2017.12.003. Epub 2017 Dec 15. Immunol Lett. 2018. PMID: 29253495
-
Injury-induced MRP8/MRP14 stimulates IP-10/CXCL10 in monocytes/macrophages.FASEB J. 2015 Jan;29(1):250-62. doi: 10.1096/fj.14-255992. Epub 2014 Oct 23. FASEB J. 2015. PMID: 25342131 Free PMC article.
-
MRP8/14 does not contribute to dissemination or inflammation in a murine model of Lyme borreliosis.Immunobiology. 2018 Nov;223(11):694-698. doi: 10.1016/j.imbio.2018.07.017. Epub 2018 Jul 17. Immunobiology. 2018. PMID: 30056999
-
Novel insights into structure and function of MRP8 (S100A8) and MRP14 (S100A9).Biochim Biophys Acta. 1998 Dec 10;1448(2):200-11. doi: 10.1016/s0167-4889(98)00144-x. Biochim Biophys Acta. 1998. PMID: 9920411 Review.
-
The calcium binding proteins MRP8 and MRP14 in acute and chronic inflammation.Behring Inst Mitt. 1992 Apr;(91):126-37. Behring Inst Mitt. 1992. PMID: 1524561 Review.
Cited by
-
Imiquimod has strain-dependent effects in mice and does not uniquely model human psoriasis.Genome Med. 2017 Mar 9;9(1):24. doi: 10.1186/s13073-017-0415-3. Genome Med. 2017. PMID: 28279190 Free PMC article.
-
Neutrophil-secreted S100A8/A9 participates in fatty liver injury and fibrosis by promoting myofibroblast migration.J Mol Med (Berl). 2024 Sep;102(9):1117-1133. doi: 10.1007/s00109-024-02469-x. Epub 2024 Jul 12. J Mol Med (Berl). 2024. PMID: 38995368
References
-
- Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. The journals of gerontology Series A, Biological sciences and medical sciences. 2014;69(Suppl 1):S4–9. - PubMed
-
- Gomes LH, Raftery MJ, Yan WX, Goyette JD, Thomas PS, Geczy CL. S100A8 and S100A9-oxidant scavengers in inflammation. Free radical biology & medicine. 2013;58:170–186. - PubMed
-
- Luo Y, Li Y, Lin C, Ma H, Zhang J, Wu S, Wang X, Gao Y, Liu Y, Zheng D. Comparative proteome analysis of splenic lymphocytes in senescence-accelerated mice. Gerontology. 2009;55:559–569. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
