

CRISPR/Cas9-Mediated Immunity to Geminiviruses: Differential Interference and Evasion
- PMID: 27225592
- PMCID: PMC4881029
- DOI: 10.1038/srep26912
CRISPR/Cas9-Mediated Immunity to Geminiviruses: Differential Interference and Evasion
Erratum in
-
Corrigendum: CRISPR/Cas9-Mediated Immunity to Geminiviruses: Differential Interference and Evasion.Sci Rep. 2016 Aug 26;6:30223. doi: 10.1038/srep30223. Sci Rep. 2016. PMID: 27561324 Free PMC article. No abstract available.
Abstract
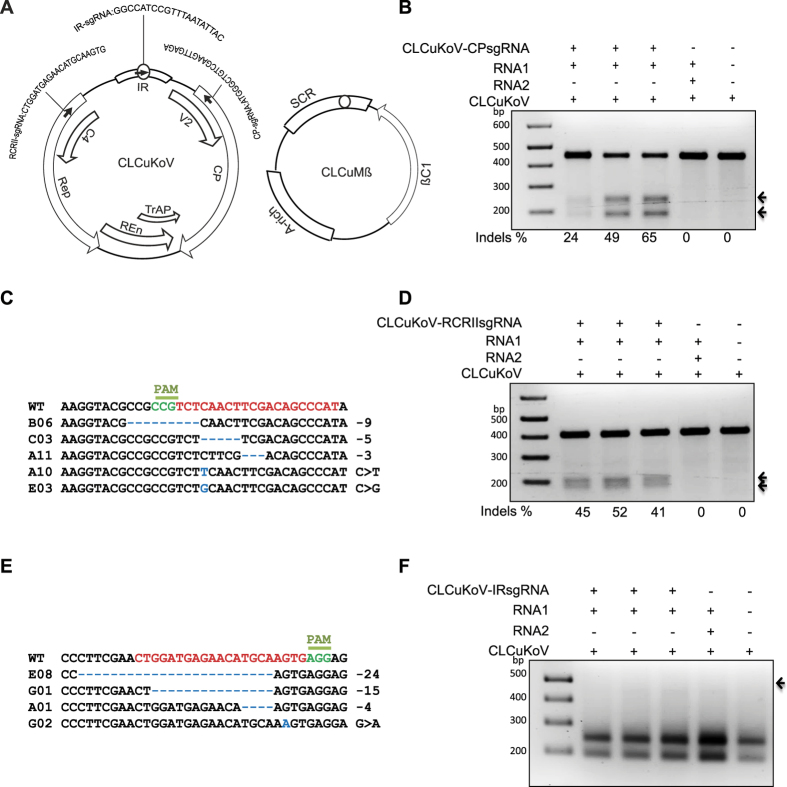
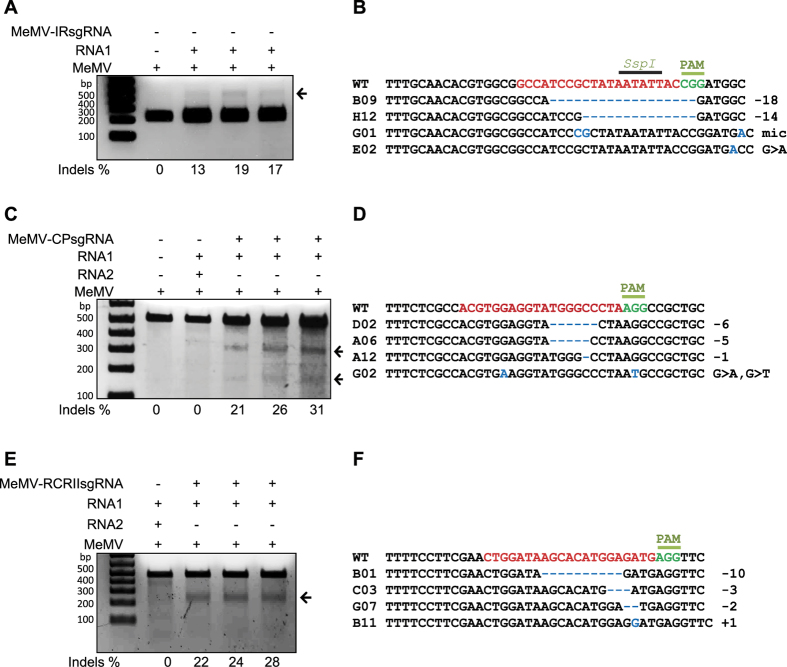
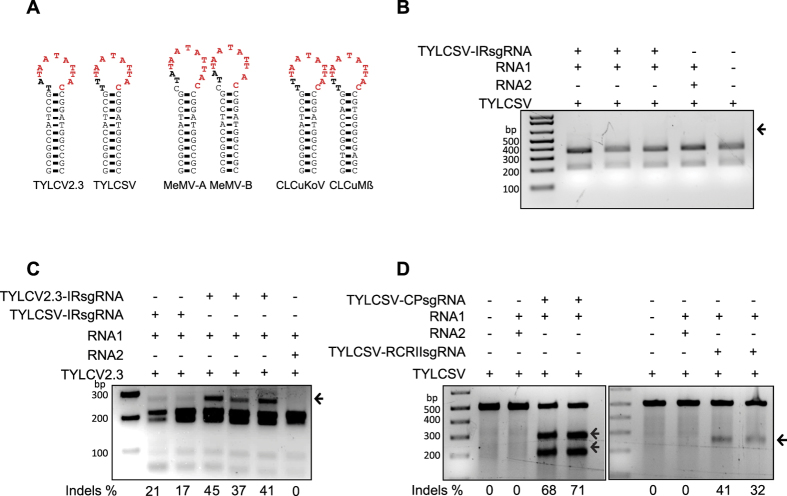
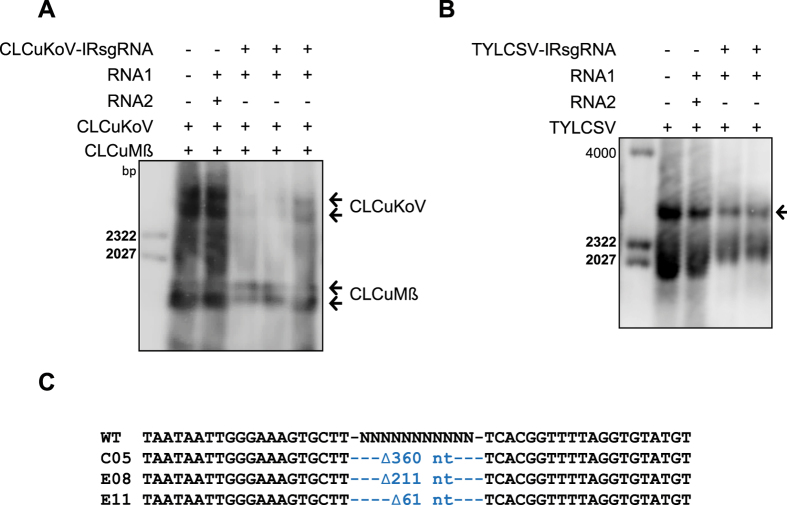
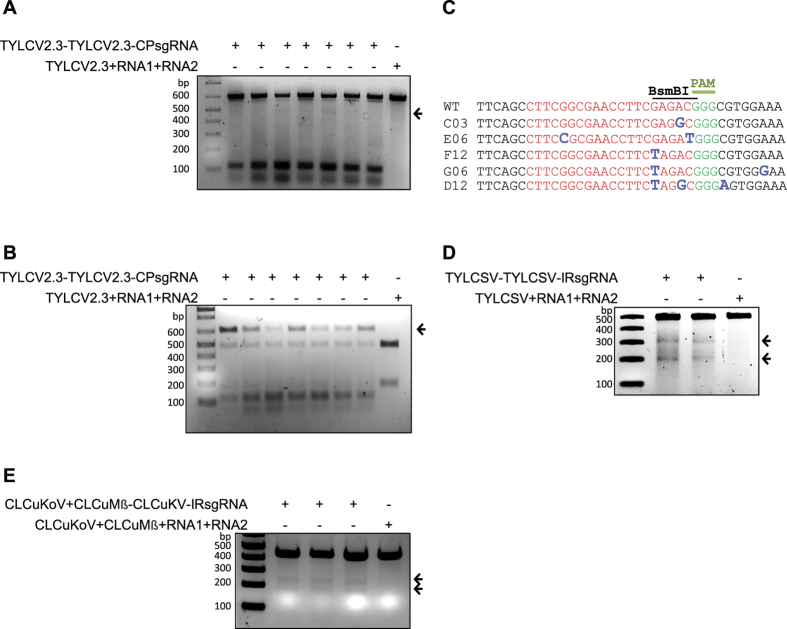
The CRISPR/Cas9 system has recently been used to confer molecular immunity against several eukaryotic viruses, including plant DNA geminiviruses. Here, we provide a detailed analysis of the efficiencies of targeting different coding and non-coding sequences in the genomes of multiple geminiviruses. Moreover, we analyze the ability of geminiviruses to evade the CRISPR/Cas9 machinery. Our results demonstrate that the CRISPR/Cas9 machinery can efficiently target coding and non-coding sequences and interfere with various geminiviruses. Furthermore, targeting the coding sequences of different geminiviruses resulted in the generation of viral variants capable of replication and systemic movement. By contrast, targeting the noncoding intergenic region sequences of geminiviruses resulted in interference, but with inefficient recovery of mutated viral variants, which thus limited the generation of variants capable of replication and movement. Taken together, our results indicate that targeting noncoding, intergenic sequences provides viral interference activity and significantly limits the generation of viral variants capable of replication and systemic infection, which is essential for developing durable resistance strategies for long-term virus control.
Conflict of interest statement
The authors declare competing financial interests. A patent application was filed on CRISPR/Cas9-mediated virus interference in plants.
Figures
References
-
- Hanley-Bowdoin L., Bejarano E. R., Robertson D. & Mansoor S. Geminiviruses: masters at redirecting and reprogramming plant processes. Nature Reviews Microbiology 11, 777–788 (2013). - PubMed
-
- Gilbertson R. L., Rojas M. & Natwick E. Development of Integrated Pest Management (IPM) Strategies for Whitefly (Bemisia tabaci)-Transmissible Geminiviruses. 323–356, 10.1007/978-94-007-1524-0_12 (2011). - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
