

Specific requirements of nonbilayer phospholipids in mitochondrial respiratory chain function and formation
- PMID: 27226479
- PMCID: PMC4945136
- DOI: 10.1091/mbc.E15-12-0865
Specific requirements of nonbilayer phospholipids in mitochondrial respiratory chain function and formation
Abstract
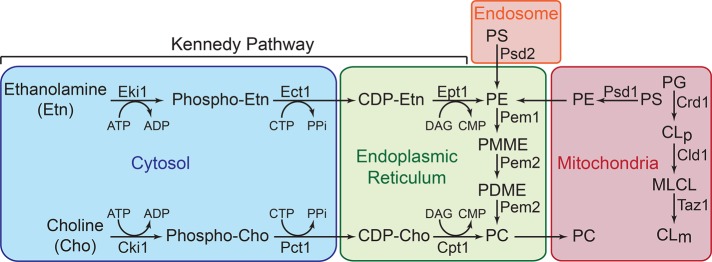
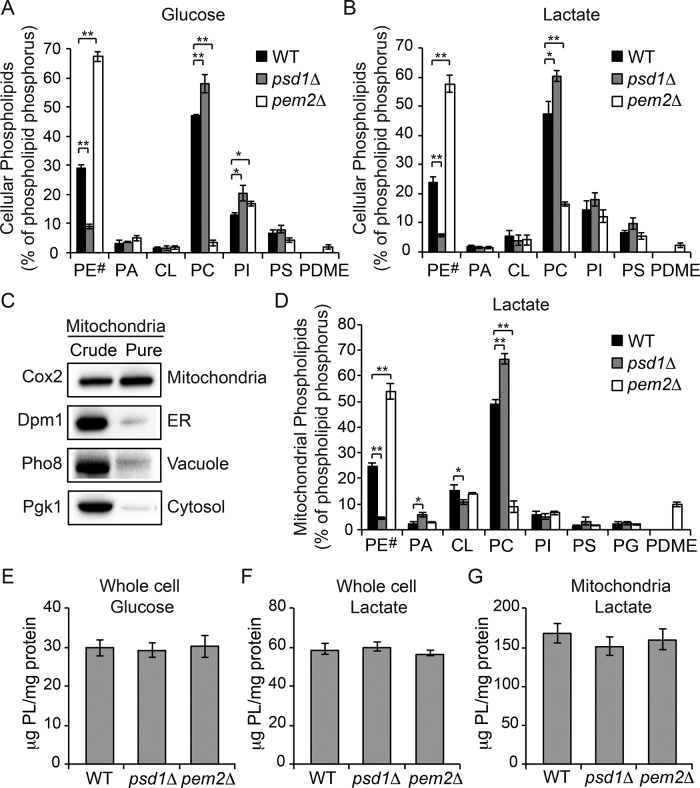
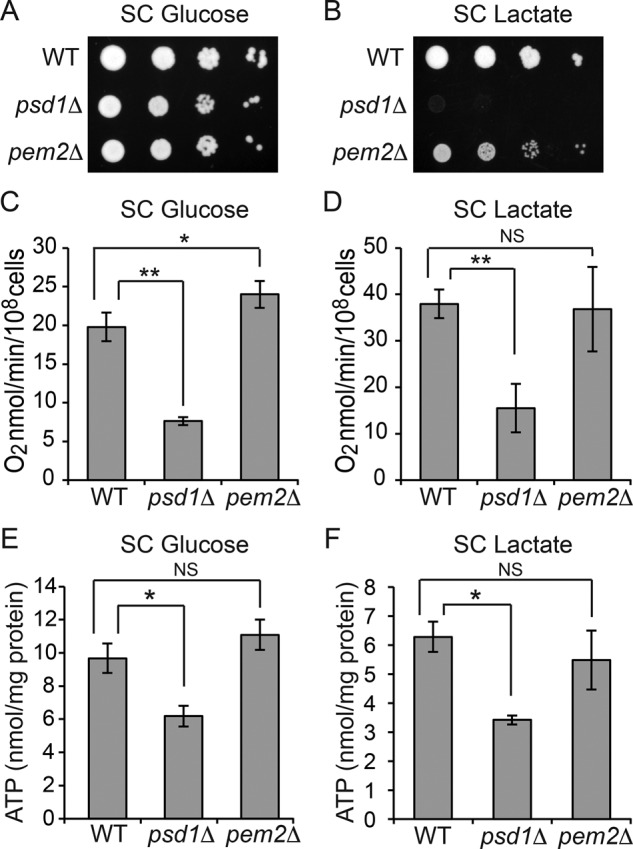
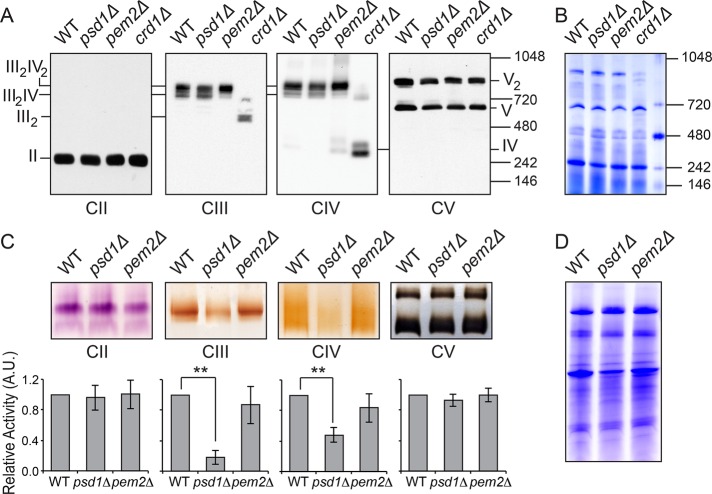
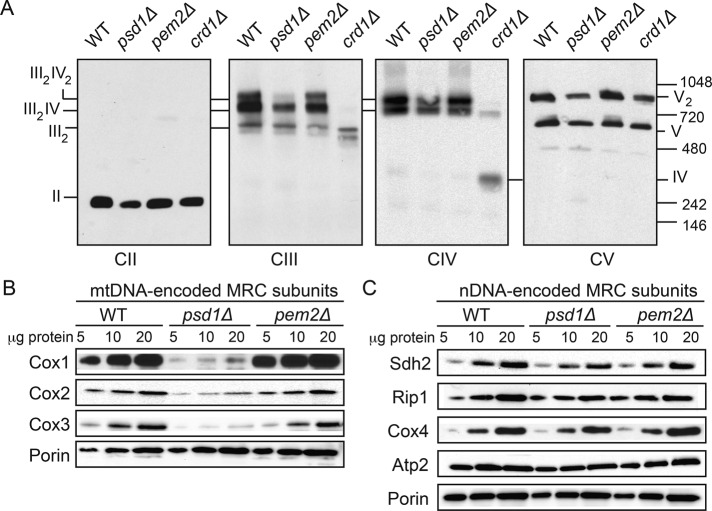
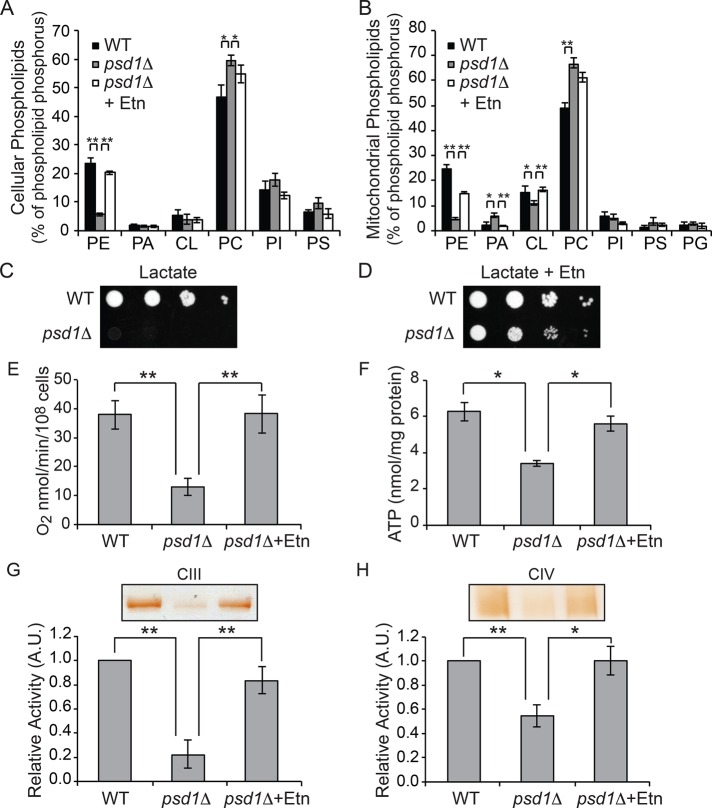
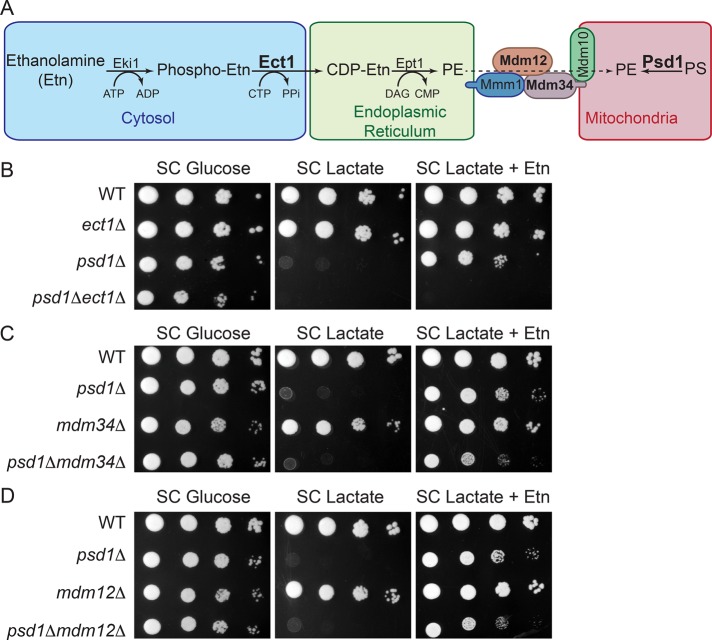
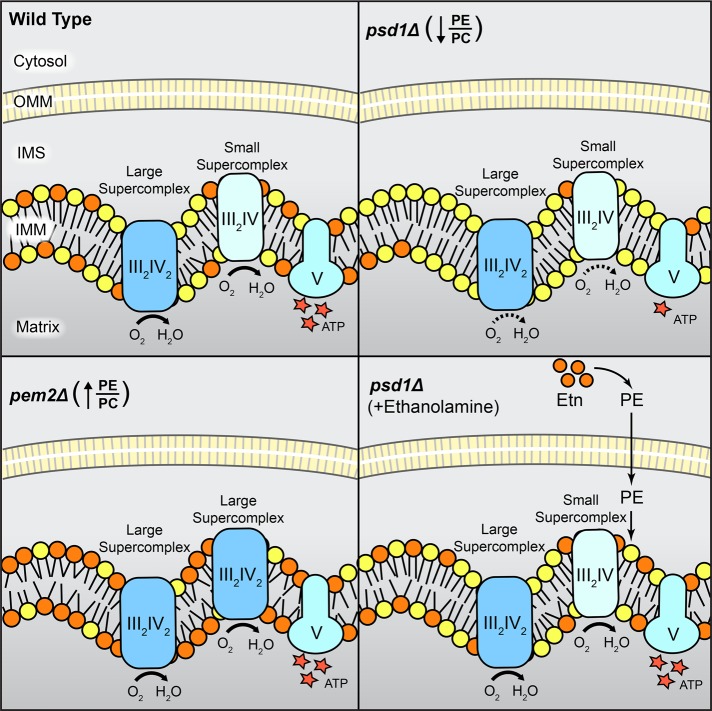
Mitochondrial membrane phospholipid composition affects mitochondrial function by influencing the assembly of the mitochondrial respiratory chain (MRC) complexes into supercomplexes. For example, the loss of cardiolipin (CL), a signature non-bilayer-forming phospholipid of mitochondria, results in disruption of MRC supercomplexes. However, the functions of the most abundant mitochondrial phospholipids, bilayer-forming phosphatidylcholine (PC) and non-bilayer-forming phosphatidylethanolamine (PE), are not clearly defined. Using yeast mutants of PE and PC biosynthetic pathways, we show a specific requirement for mitochondrial PE in MRC complex III and IV activities but not for their formation, whereas loss of PC does not affect MRC function or formation. Unlike CL, mitochondrial PE or PC is not required for MRC supercomplex formation, emphasizing the specific requirement of CL in supercomplex assembly. Of interest, PE biosynthesized in the endoplasmic reticulum (ER) can functionally substitute for the lack of mitochondrial PE biosynthesis, suggesting the existence of PE transport pathway from ER to mitochondria. To understand the mechanism of PE transport, we disrupted ER-mitochondrial contact sites formed by the ERMES complex and found that, although not essential for PE transport, ERMES facilitates the efficient rescue of mitochondrial PE deficiency. Our work highlights specific roles of non-bilayer-forming phospholipids in MRC function and formation.
© 2016 Baker et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Acín-Pérez R, Fernández-Silva P, Peleato ML, Pérez-Martos A, Enriquez JA. Respiratory active mitochondrial supercomplexes. Mol Cell. 2008;32:529–539. - PubMed
-
- Aktas M, Wessel M, Hacker S, Klusener S, Gleichenhagen J, Narberhaus F. Phosphatidylcholine biosynthesis and its significance in bacteria interacting with eukaryotic cells. Eur J Cell Biol. 2010;89:888–894. - PubMed
-
- Amberg DC, Burke DJ, Strathern JN. Methods in Yeast Genetics. A Cold Spring Harbor Laboratory Course Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2005.
-
- Bartlett GR. Phosphorus assay in column chromatography. J Biol Chem. 1959;234:466–468. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
