

S100A1 Protein Does Not Compete with Calmodulin for Ryanodine Receptor Binding but Structurally Alters the Ryanodine Receptor·Calmodulin Complex
- PMID: 27226555
- PMCID: PMC4957069
- DOI: 10.1074/jbc.M115.713107
S100A1 Protein Does Not Compete with Calmodulin for Ryanodine Receptor Binding but Structurally Alters the Ryanodine Receptor·Calmodulin Complex
Abstract
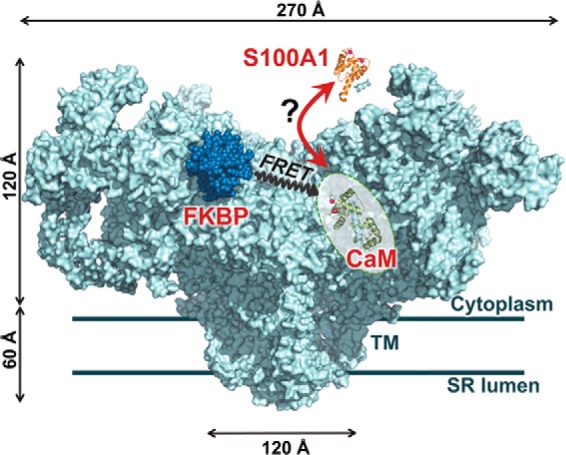
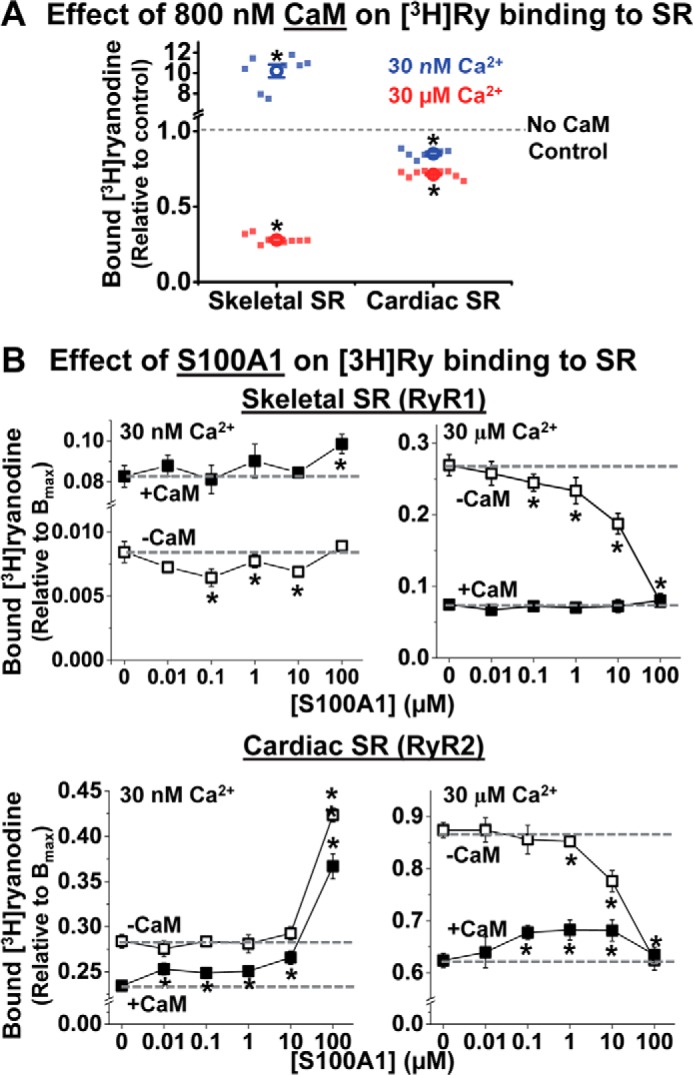
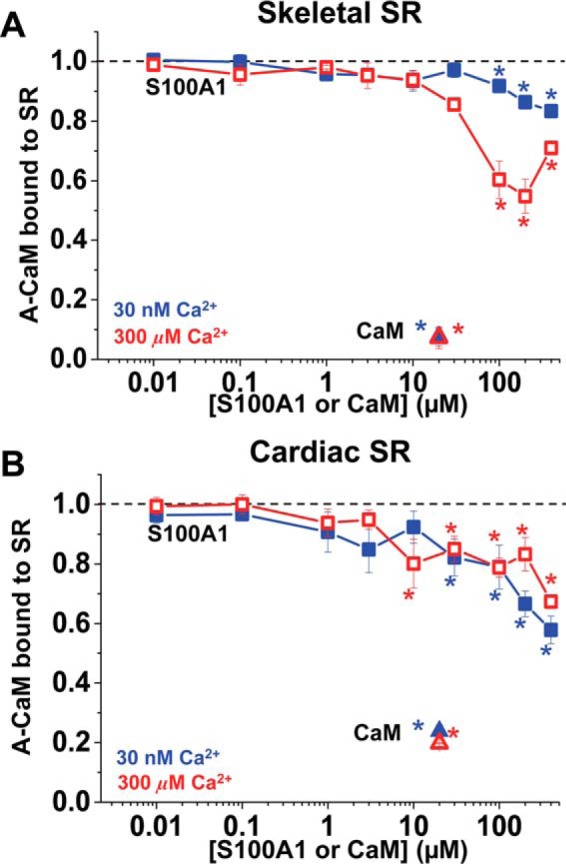
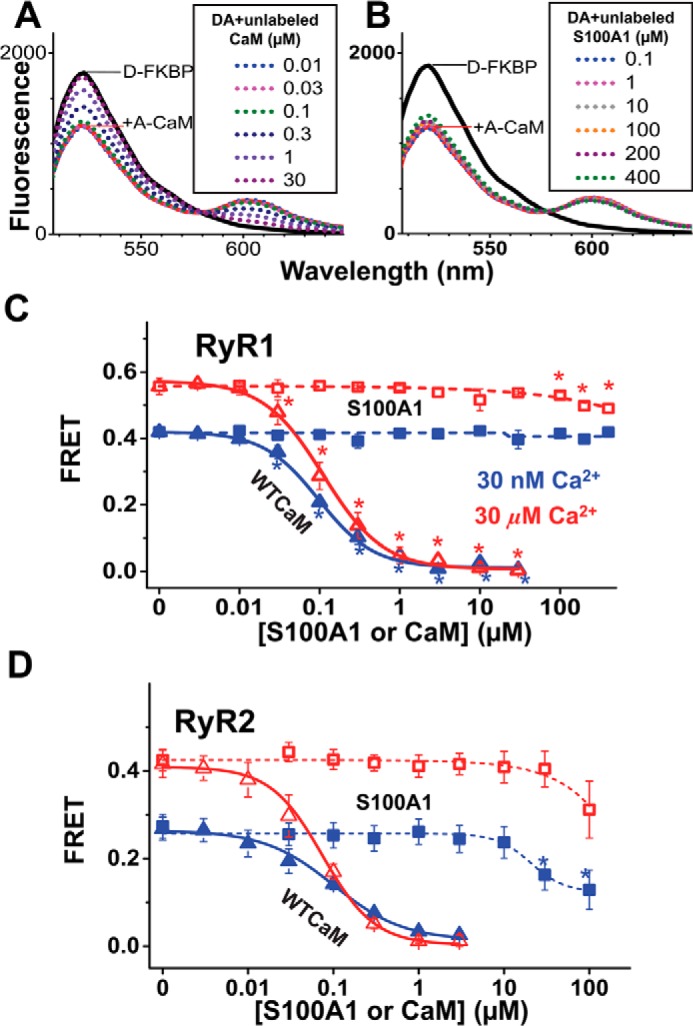
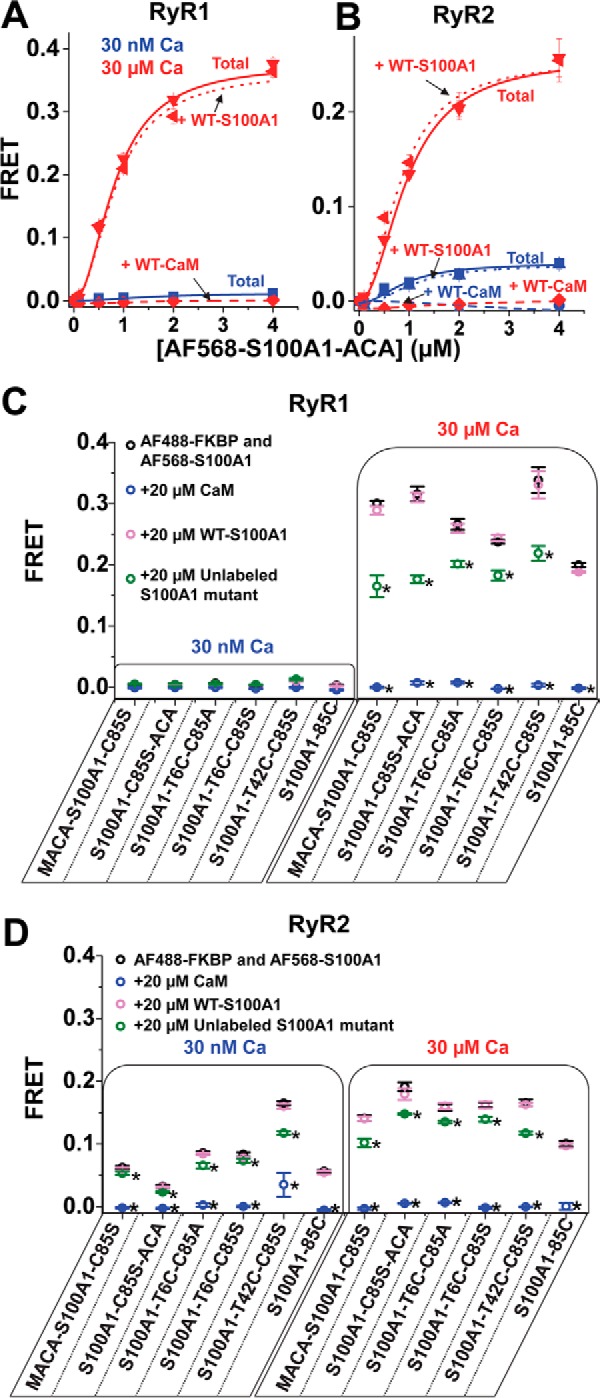
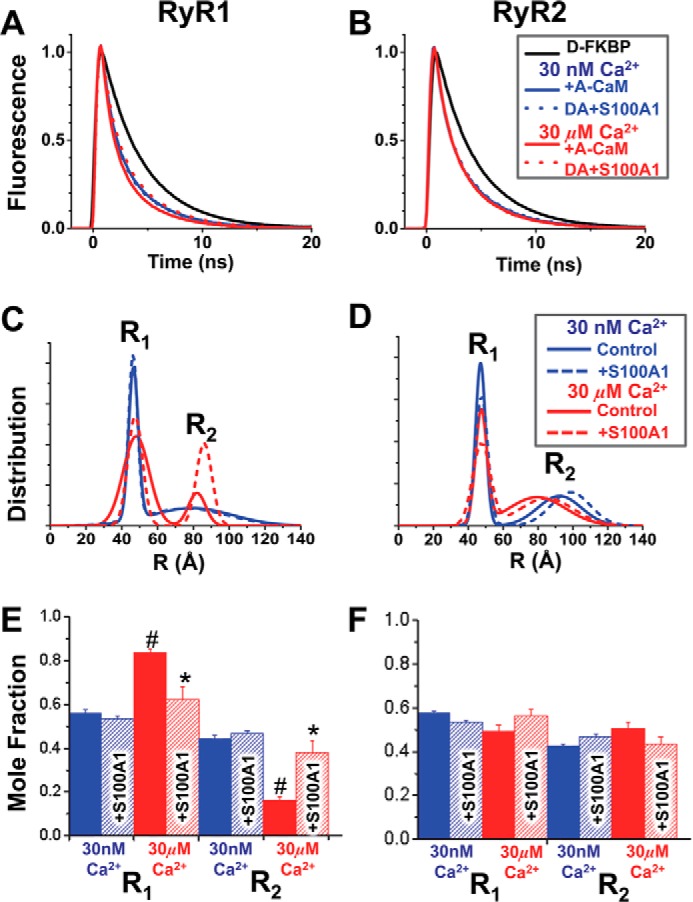
S100A1 has been suggested as a therapeutic agent to enhance myocyte Ca(2+) cycling in heart failure, but its molecular mode of action is poorly understood. Using FRET, we tested the hypothesis that S100A1 directly competes with calmodulin (CaM) for binding to intact, functional ryanodine receptors type I (RyR1) and II (RyR2) from skeletal and cardiac muscle, respectively. Our FRET readout provides an index of acceptor-labeled CaM binding near donor-labeled FKBP (FK506-binding protein 12.6) on the cytoplasmic domain of RyR in isolated sarcoplasmic reticulum vesicles. S100A1 (0.01-400 μm) partially inhibited FRET (i.e. CaM binding), with Ki > 10 μm, for both RyR1 and RyR2. The high [S100A1] required for partial effects on FRET indicates a lack of competition by S100A1 on CaM/RyR binding under normal physiological conditions. High-resolution analysis of time-resolved FRET detects two structural states of RyR-bound CaM, which respond to [Ca(2+)] and are isoform-specific. The distribution of these structural states was perturbed only by high micromolar [S100A1], which promoted a shift of bound CaM to a lower FRET orientation (without altering the amount of CaM bound to RyR). Thus, high micromolar S100A1 does alter the CaM/RyR interaction, without involving competition. Nevertheless, submicromolar S100A1 can alter RyR function, an effect that is influenced by both [Ca(2+)] and [CaM]. We conclude that CaM and S100A1 can concurrently bind to and functionally modulate RyR1 and RyR2, but this does not involve direct competition at the RyR CaM binding site.
Keywords: FKBP12.6; calcium channel; calmodulin (CaM); fluorescence lifetime; fluorescence resonance energy transfer (FRET); sarcoplasmic reticulum (SR); structure-function.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Buratti R., Prestipino G., Menegazzi P., Treves S., and Zorzato F. (1995) Calcium dependent activation of skeletal muscle Ca2+ release channel (ryanodine receptor) by calmodulin. Biochem. Biophys. Res. Commun. 213, 1082–1090 - PubMed
-
- Treves S., Scutari E., Robert M., Groh S., Ottolia M., Prestipino G., Ronjat M., and Zorzato F. (1997) Interaction of S100A1 with the Ca2+ release channel (ryanodine receptor) of skeletal muscle. Biochemistry 36, 11496–11503 - PubMed
-
- Kettlewell S., Most P., Currie S., Koch W. J., and Smith G. L. (2005) S100A1 increases the gain of excitation-contraction coupling in isolated rabbit ventricular cardiomyocytes. J. Mol. Cell Cardiol. 39, 900–910 - PubMed
-
- Völkers M., Loughrey C. M., Macquaide N., Remppis A., DeGeorge B. R. Jr., Wegner F. V., Friedrich O., Fink R. H., Koch W. J., Smith G. L., and Most P. (2007) S100A1 decreases calcium spark frequency and alters their spatial characteristics in permeabilized adult ventricular cardiomyocytes. Cell Calcium 41, 135–143 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
