

Heterogeneity in Cancer Metabolism: New Concepts in an Old Field
- PMID: 27228792
- PMCID: PMC5359687
- DOI: 10.1089/ars.2016.6750
Heterogeneity in Cancer Metabolism: New Concepts in an Old Field
Abstract
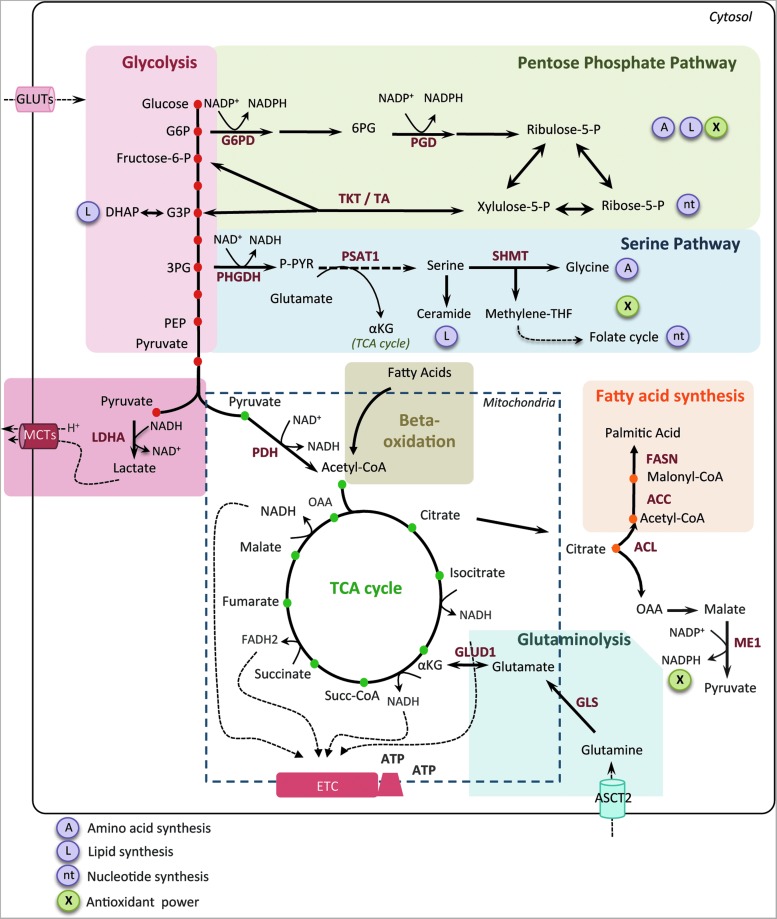
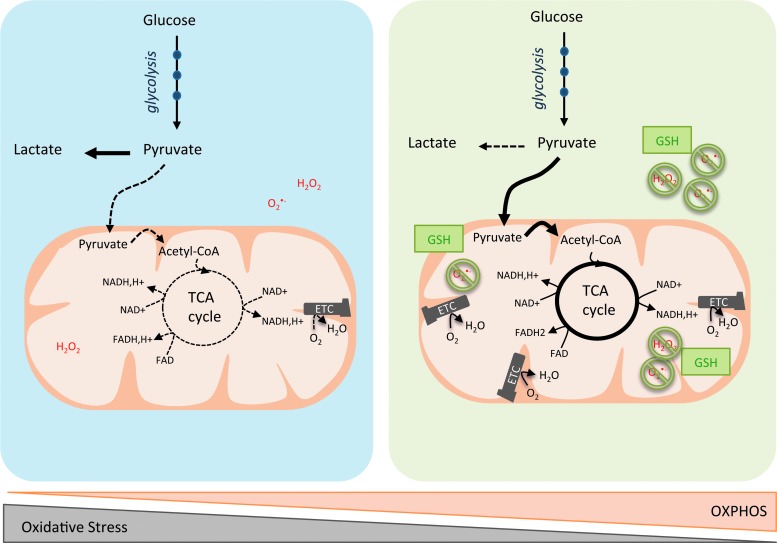
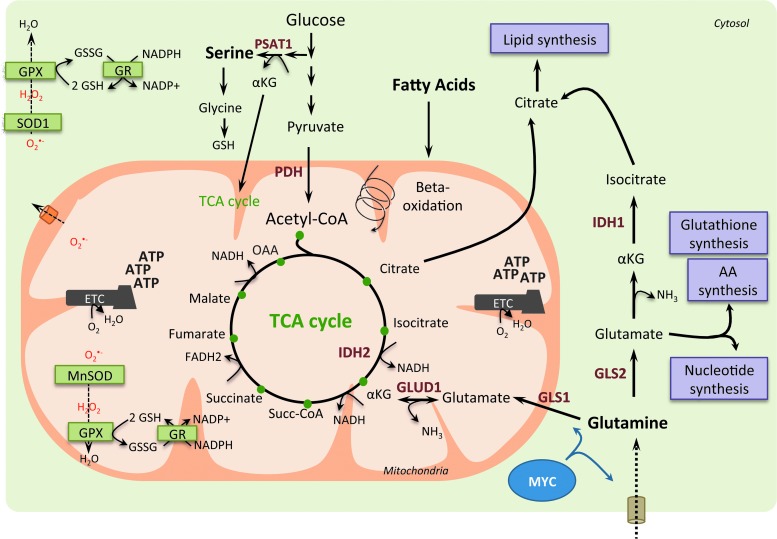
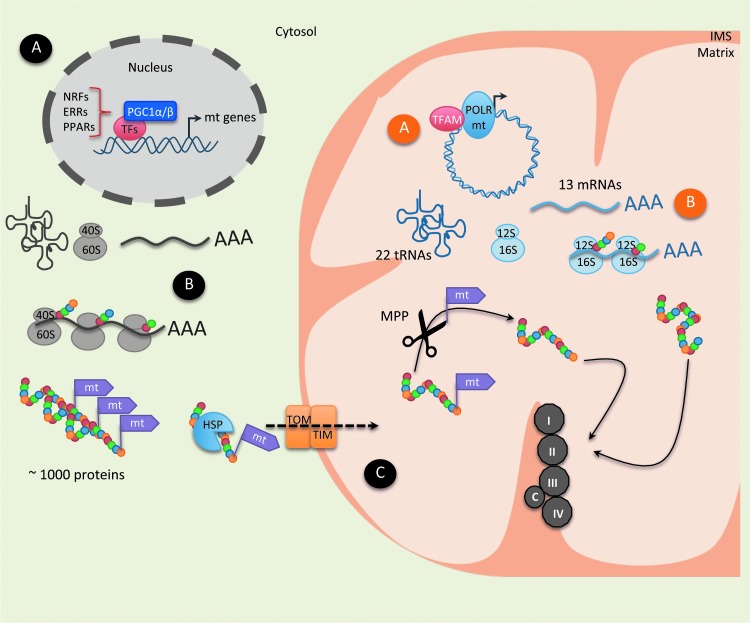
Significance: In the last years, metabolic reprogramming, fluctuations in bioenergetic fuels, and modulation of oxidative stress became new key hallmarks of tumor development. In cancer, elevated glucose uptake and high glycolytic rate, as a source of adenosine triphosphate, constitute a growth advantage for tumors. This represents the universally known Warburg effect, which gave rise to one major clinical application for detecting cancer cells using glucose analogs: the positron emission tomography scan imaging. Recent Advances: Glucose utilization and carbon sources in tumors are much more heterogeneous than initially thought. Indeed, new studies emerged and revealed a dual capacity of tumor cells for glycolytic and oxidative phosphorylation (OXPHOS) metabolism. OXPHOS metabolism, which relies predominantly on mitochondrial respiration, exhibits fine-tuned regulation of respiratory chain complexes and enhanced antioxidant response or detoxification capacity.
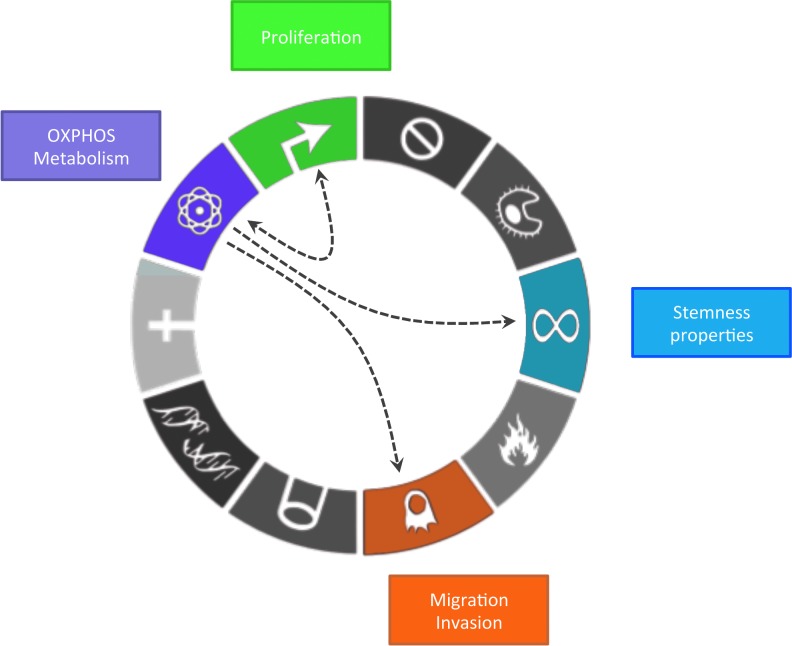
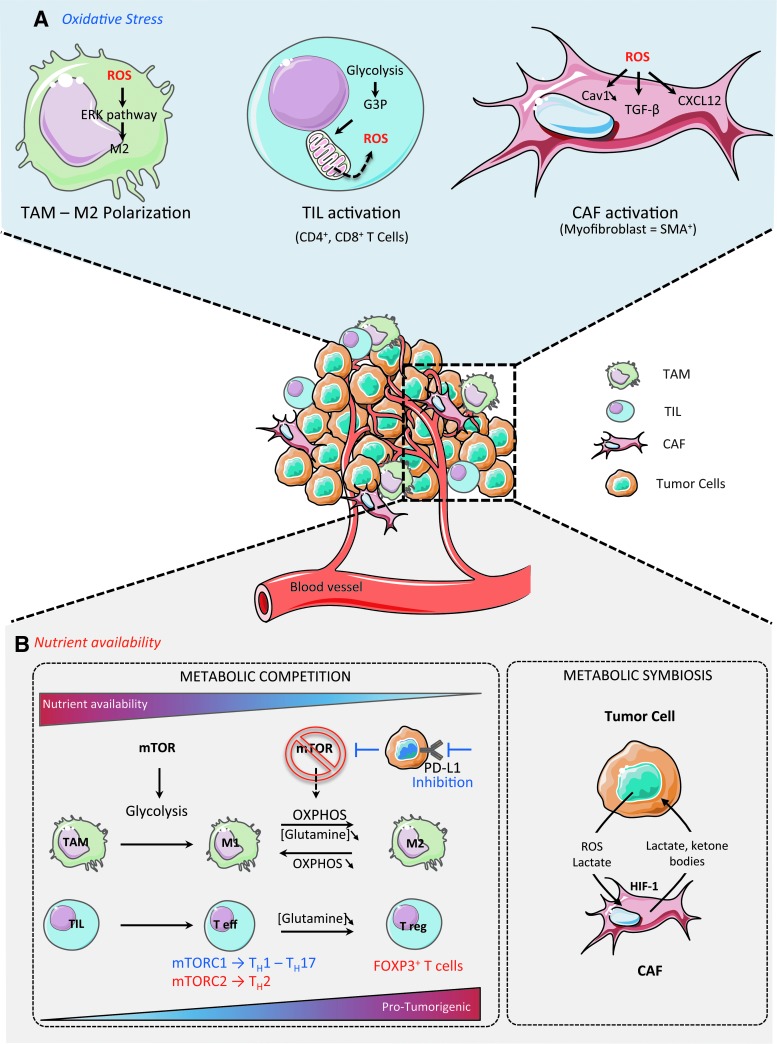
Critical issues: OXPHOS-dependent cancer cells use alternative oxidizable substrates, such as glutamine and fatty acids. The diversity of carbon substrates fueling neoplastic cells is indicative of metabolic heterogeneity, even within tumors sharing the same clinical diagnosis. Metabolic switch supports cancer cell stemness and their bioenergy-consuming functions, such as proliferation, survival, migration, and invasion. Moreover, reactive oxygen species-induced mitochondrial metabolism and nutrient availability are important for interaction with tumor microenvironment components. Carcinoma-associated fibroblasts and immune cells participate in the metabolic interplay with neoplastic cells. They collectively adapt in a dynamic manner to the metabolic needs of cancer cells, thus participating in tumorigenesis and resistance to treatments.
Future directions: Characterizing the reciprocal metabolic interplay between stromal, immune, and neoplastic cells will provide a better understanding of treatment resistance. Antioxid. Redox Signal. 26, 462-485.
Keywords: ROS; cancer; immunology; metabolism; mitochondria; oxidative stress.
Figures
References
-
- Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F, Schreier PH, Smith AJ, Staden R, and Young IG. Sequence and organization of the human mitochondrial genome. Nature 290: 457–465, 1981 - PubMed
-
- Antoch G, Saoudi N, Kuehl H, Dahmen G, Mueller SP, Beyer T, Bockisch A, Debatin JF, and Freudenberg LS. Accuracy of whole-body dual-modality fluorine-18–2-fluoro-2-deoxy-D-glucose positron emission tomography and computed tomography (FDG-PET/CT) for tumor staging in solid tumors: comparison with CT and PET. J Clin Oncol 22: 4357–4368, 2004 - PubMed
-
- Augsten M, Sjoberg E, Frings O, Vorrink SU, Frijhoff J, Olsson E, Borg A, and Ostman A. Cancer-associated fibroblasts expressing CXCL14 rely upon NOS1-derived nitric oxide signaling for their tumor-supporting properties. Cancer Res 74: 2999–3010, 2014 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
