

Pre-bilaterian origin of the blastoporal axial organizer
- PMID: 27229764
- PMCID: PMC4895019
- DOI: 10.1038/ncomms11694
Pre-bilaterian origin of the blastoporal axial organizer
Abstract
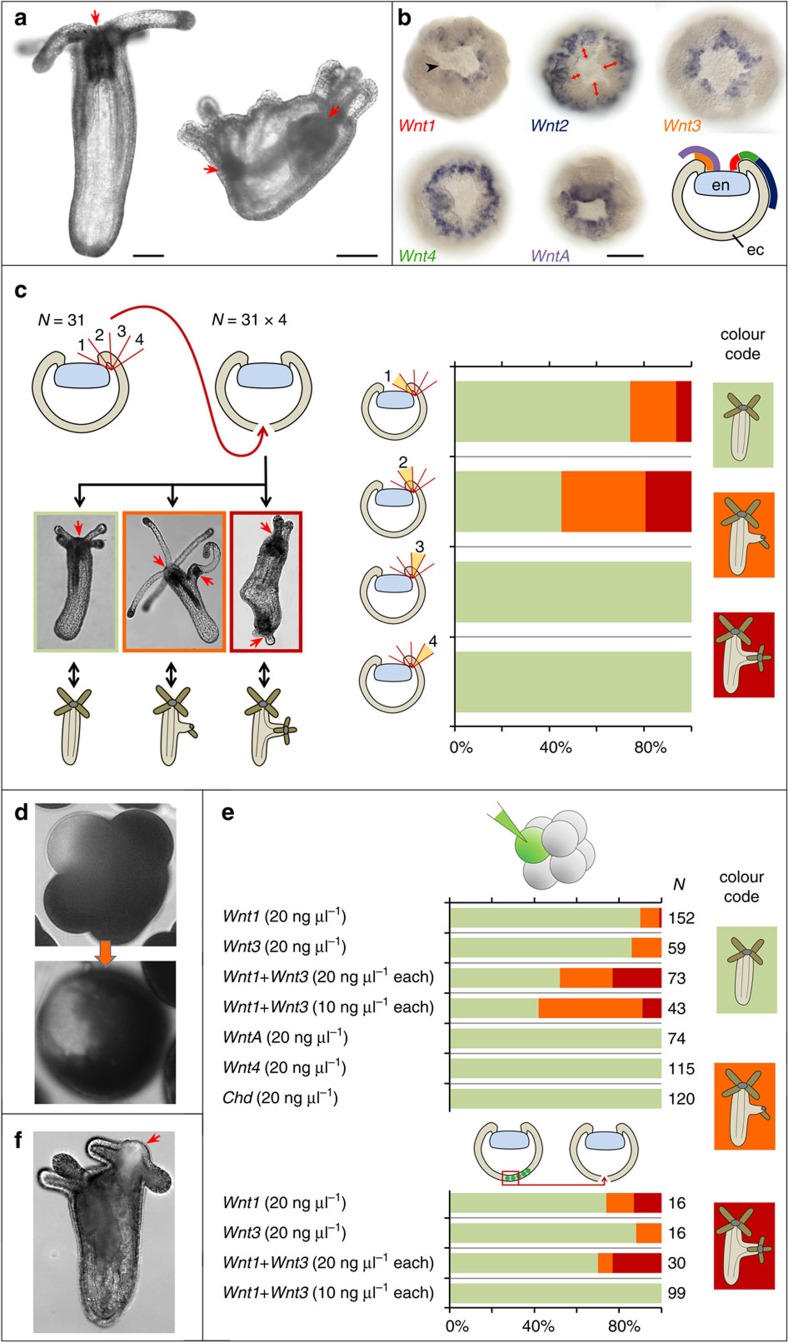
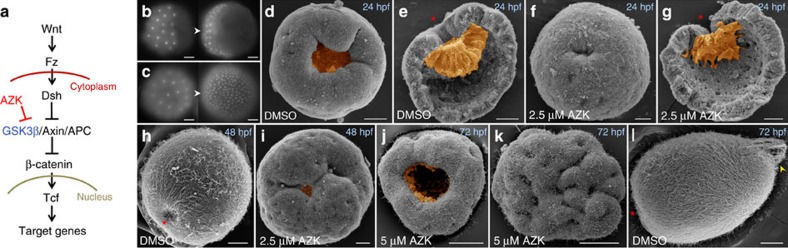
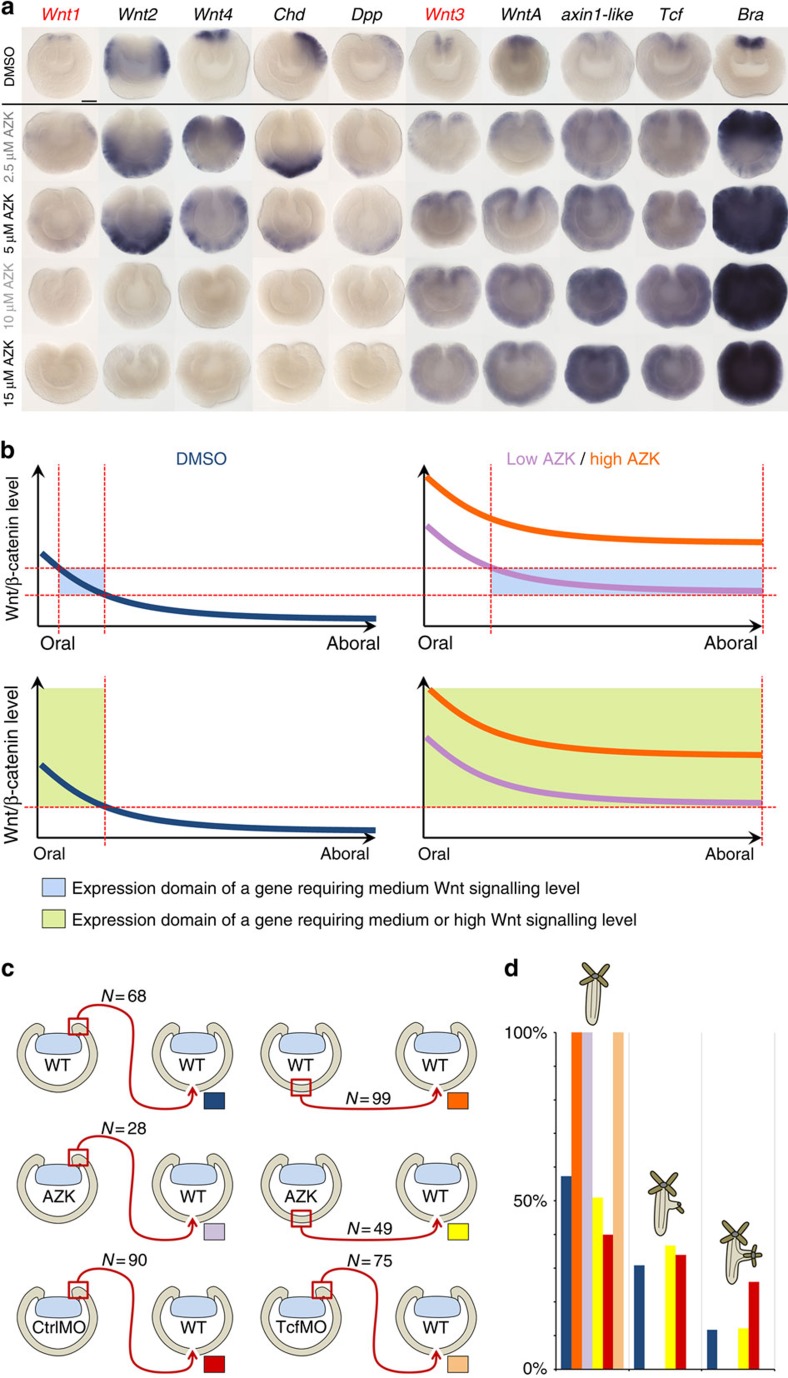
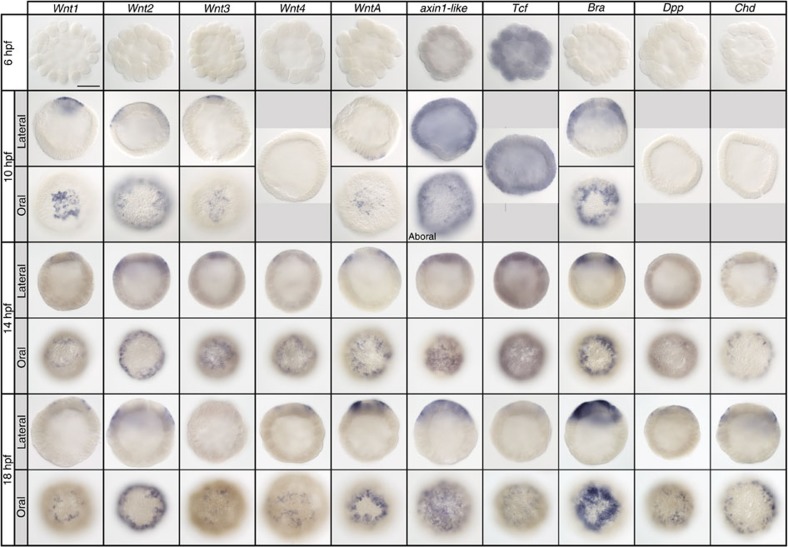
The startling capacity of the amphibian Spemann organizer to induce naïve cells to form a Siamese twin embryo with a second set of body axes is one of the hallmarks of developmental biology. However, the axis-inducing potential of the blastopore-associated tissue is commonly regarded as a chordate feature. Here we show that the blastopore lip of a non-bilaterian metazoan, the anthozoan cnidarian Nematostella vectensis, possesses the same capacity and uses the same molecular mechanism for inducing extra axes as chordates: Wnt/β-catenin signaling. We also demonstrate that the establishment of the secondary, directive axis in Nematostella by BMP signaling is sensitive to an initial Wnt signal, but once established the directive axis becomes Wnt-independent. By combining molecular analysis with experimental embryology, we provide evidence that the emergence of the Wnt/β-catenin driven blastopore-associated axial organizer predated the cnidarian-bilaterian split over 600 million years ago.
Figures
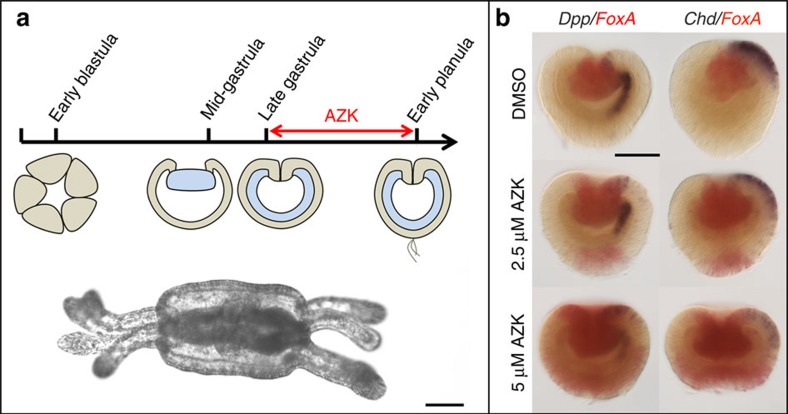
References
-
- Spemann H. & Mangold H. Über Induktion von Embryonalanlagen durch Implantation artfremder Organisatoren. W. Roux's Arch. Entwicklungsmech. Organ 100, 599–638 (1924).
-
- Kraus Y., Fritzenwanker J. H., Genikhovich G. & Technau U. The blastoporal organiser of a sea anemone. Curr. Biol. 17, R874–R876 (2007). - PubMed
-
- Bouwmeester T., Kim S., Sasai Y., Lu B. & De Robertis E. M. Cerberus is a head-inducing secreted factor expressed in the anterior endoderm of Spemann's organizer. Nature 382, 595–601 (1996). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
