

Neutrophil migration in infection and wound repair: going forward in reverse
- PMID: 27231052
- PMCID: PMC5367630
- DOI: 10.1038/nri.2016.49
Neutrophil migration in infection and wound repair: going forward in reverse
Abstract
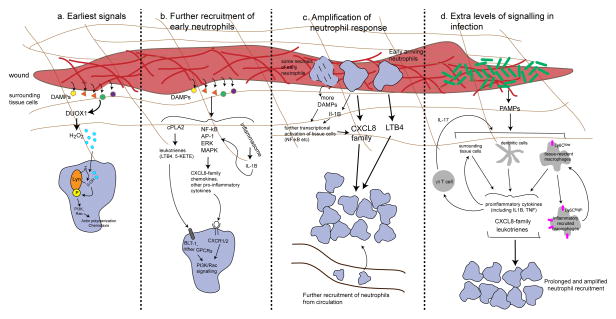
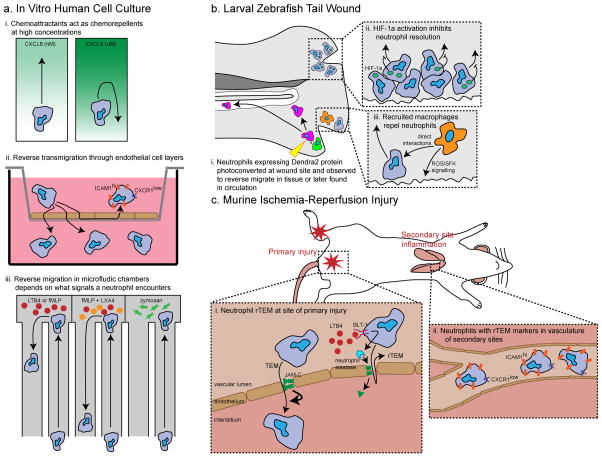
Neutrophil migration and its role during inflammation has been the focus of increased interest in the past decade. Advances in live imaging and the use of new model systems have helped to uncover the behaviour of neutrophils in injured and infected tissues. Although neutrophils were considered to be short-lived effector cells that undergo apoptosis in damaged tissues, recent evidence suggests that neutrophil behaviour is more complex and, in some settings, neutrophils might leave sites of tissue injury and migrate back into the vasculature. The role of reverse migration and its contribution to resolution of inflammation remains unclear. In this Review, we discuss the different cues within tissues that mediate neutrophil forward and reverse migration in response to injury or infection and the implications of these mechanisms to human disease.
Figures
References
-
- Kolaczkowska E, Kubes P. Neutrophil recruitment and function in health and inflammation. Nature reviews. Immunology. 2013;13:159–175. - PubMed
-
- Nathan C. Neutrophils and immunity: challenges and opportunities. Nature reviews. Immunology. 2006;6:173–182. - PubMed
-
- Borregaard N. Neutrophils, from marrow to microbes. Immunity. 2010;33:657–670. - PubMed
-
- Ley K. Integration of inflammatory signals by rolling neutrophils. Immunol Rev. 2002;186:8–18. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
