

Thermal stability of human plasma electronegative low-density lipoprotein: A paradoxical behavior of low-density lipoprotein aggregation
- PMID: 27233433
- PMCID: PMC5572826
- DOI: 10.1016/j.bbalip.2016.05.008
Thermal stability of human plasma electronegative low-density lipoprotein: A paradoxical behavior of low-density lipoprotein aggregation
Abstract
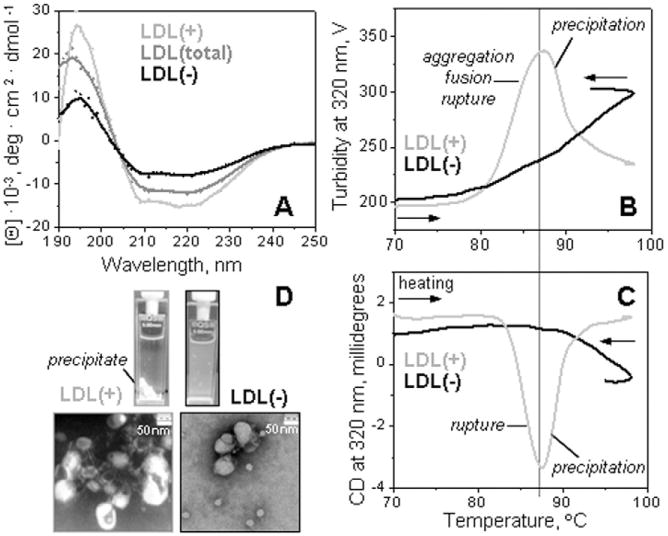
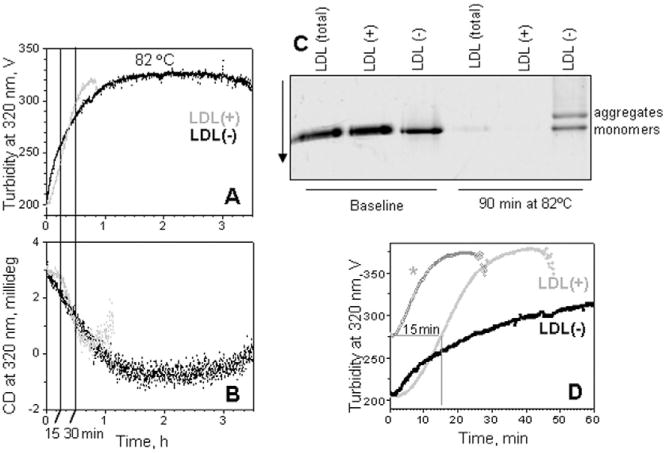
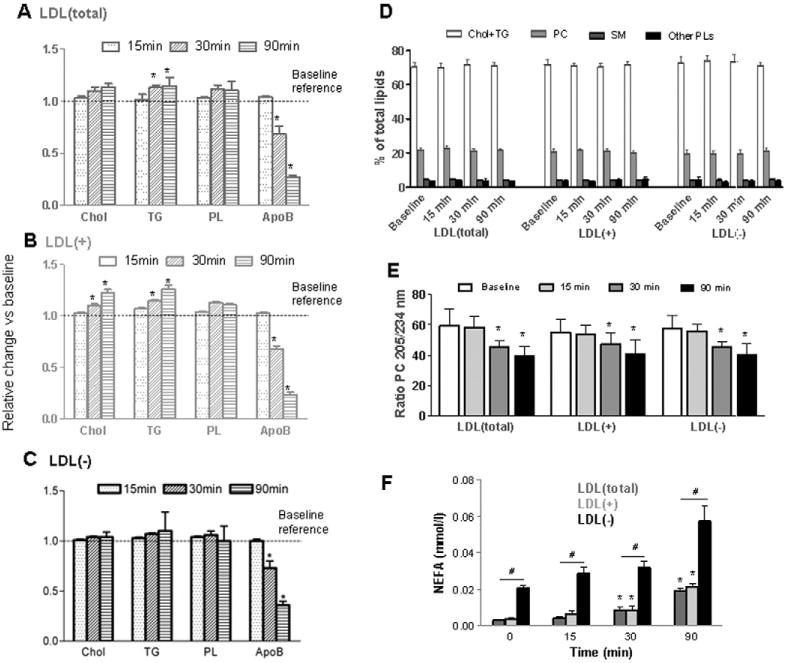
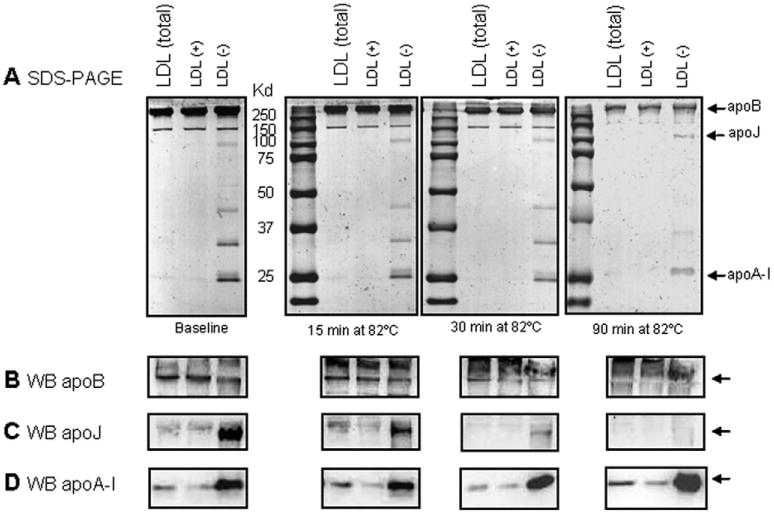
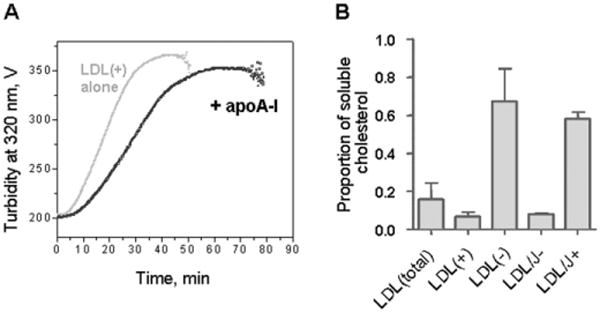
Low-density lipoprotein (LDL) aggregation is central in triggering atherogenesis. A minor fraction of electronegative plasma LDL, termed LDL(-), plays a special role in atherogenesis. To better understand this role, we analyzed the kinetics of aggregation, fusion and disintegration of human LDL and its fractions, LDL(+) and LDL(-). Thermal denaturation of LDL was monitored by spectroscopy and electron microscopy. Initially, LDL(-) aggregated and fused faster than LDL(+), but later the order reversed. Most LDL(+) disintegrated and precipitated upon prolonged heating. In contrast, LDL(-) partially retained lipoprotein morphology and formed soluble aggregates. Biochemical analysis of all fractions showed no significant degradation of major lipids, mild phospholipid oxidation, and an increase in non-esterified fatty acid (NEFA) upon thermal denaturation. The main baseline difference between LDL subfractions was higher content of NEFA in LDL(-). Since NEFA promote lipoprotein fusion, increased NEFA content can explain rapid initial aggregation and fusion of LDL(-) but not its resistance to extensive disintegration. Partial hydrolysis of apoB upon heating was similar in LDL subfractions, suggesting that minor proteins importantly modulate LDL disintegration. Unlike LDL(+), LDL(-) contains small amounts of apoA-I and apoJ. Addition of exogenous apoA-I to LDL(+) hampered lipoprotein aggregation, fusion and precipitation, while depletion of endogenous apoJ had an opposite effect. Therefore, the initial rapid aggregation of LDL(-) is apparently counterbalanced by the stabilizing effects of minor proteins such as apoA-I and apoJ. These results help identify key determinants for LDL aggregation, fusion and coalescence into lipid droplets in vivo.
Keywords: Apolipoprotein A-I; Apolipoprotein J; Atherogenesis; Electronegative LDL; Lipoprotein aggregation, fusion and droplet formation; Thermal denaturation.
Copyright © 2016 Elsevier B.V. All rights reserved.
Figures

Similar articles
-
Clusterin/apolipoprotein J binds to aggregated LDL in human plasma and plays a protective role against LDL aggregation.FASEB J. 2015 May;29(5):1688-700. doi: 10.1096/fj.14-264036. Epub 2014 Dec 30. FASEB J. 2015. PMID: 25550461
-
Aggregated electronegative low density lipoprotein in human plasma shows a high tendency toward phospholipolysis and particle fusion.J Biol Chem. 2010 Oct 15;285(42):32425-35. doi: 10.1074/jbc.M110.139691. Epub 2010 Jul 29. J Biol Chem. 2010. PMID: 20670941 Free PMC article.
-
Low-density lipoprotein aggregation is inhibited by apolipoprotein J-derived mimetic peptide D-[113-122]apoJ.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Feb;1865(2):158541. doi: 10.1016/j.bbalip.2019.158541. Epub 2019 Oct 28. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 31672573
-
Electronegative low-density lipoprotein. A link between apolipoprotein B misfolding, lipoprotein aggregation and proteoglycan binding.Curr Opin Lipidol. 2012 Oct;23(5):479-86. doi: 10.1097/MOL.0b013e328357c933. Curr Opin Lipidol. 2012. PMID: 22964994 Review.
-
Circulating low density lipoprotein (LDL).Horm Mol Biol Clin Investig. 2018 Jul 31;35(2):/j/hmbci.2018.35.issue-2/hmbci-2018-0024/hmbci-2018-0024.xml. doi: 10.1515/hmbci-2018-0024. Horm Mol Biol Clin Investig. 2018. PMID: 30059347 Review.
Cited by
-
Effects of triacylglycerol on the structural remodeling of human plasma very low- and low-density lipoproteins.Biochim Biophys Acta Mol Cell Biol Lipids. 2019 Jul;1864(7):1061-1071. doi: 10.1016/j.bbalip.2019.03.001. Epub 2019 Mar 5. Biochim Biophys Acta Mol Cell Biol Lipids. 2019. PMID: 30844432 Free PMC article.
-
We Must Abandon the Myth: Oxidized Low-density Lipoprotein is not a Lipoprotein that Plays a Key Role in Atherogenesis.Curr Med Chem. 2025;32(15):2899-2914. doi: 10.2174/0109298673301236240311113807. Curr Med Chem. 2025. PMID: 38494931 Free PMC article. Review.
-
LDL binding to cell receptors and extracellular matrix is proatherogenic in obesity but improves after bariatric surgery.J Lipid Res. 2023 Nov;64(11):100451. doi: 10.1016/j.jlr.2023.100451. Epub 2023 Sep 28. J Lipid Res. 2023. PMID: 37777014 Free PMC article.
-
Triglyceride increase in the core of high-density lipoproteins augments apolipoprotein dissociation from the surface: Potential implications for treatment of apolipoprotein deposition diseases.Biochim Biophys Acta Mol Basis Dis. 2017 Jan;1863(1):200-210. doi: 10.1016/j.bbadis.2016.10.010. Epub 2016 Oct 18. Biochim Biophys Acta Mol Basis Dis. 2017. PMID: 27768903 Free PMC article.
-
Binding to heparin triggers deleterious structural and biochemical changes in human low-density lipoprotein, which are amplified in hyperglycemia.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Aug;1865(8):158712. doi: 10.1016/j.bbalip.2020.158712. Epub 2020 Apr 11. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 32289504 Free PMC article.
References
-
- Williams KJ, Tabas I. The response-to-retention hypothesis of atherogenesis reinforced. Curr Opin Lipidol. 1998;9:471–474. - PubMed
-
- Williams KJ, Tabas I. Lipoprotein retention – and clues for atheroma regression. Arterioscler Thromb Vasc Biol. 2005;25:1536–1540. - PubMed
-
- Oorni K, Pentikainen MO, Ala-Korpela M, Kovanen PT. Aggregation, fusion, and vesicle formation of modified low density lipoprotein particles: molecular mechanisms and effects on matrix interactions. J Lipid Res. 2000;41:1703–1714. - PubMed
-
- Pentikainen MO, Oorni K, Ala-Korpela M, Kovanen PT. Modified LDL - trigger of atherosclerosis and inflammation in the arterial intima. J Intern Med. 2000;247:359–370. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
