

Calcium dynamics predict direction of synaptic plasticity in striatal spiny projection neurons
- PMID: 27233469
- PMCID: PMC5124545
- DOI: 10.1111/ejn.13287
Calcium dynamics predict direction of synaptic plasticity in striatal spiny projection neurons
Abstract
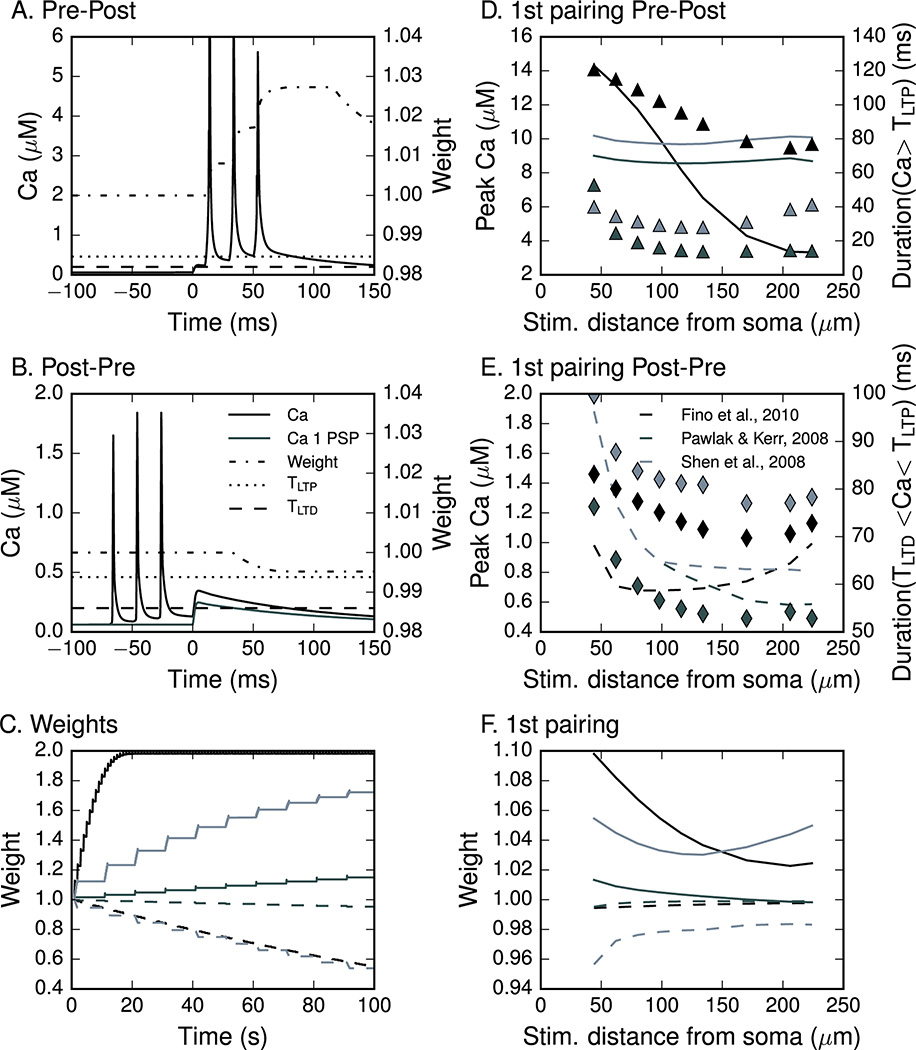
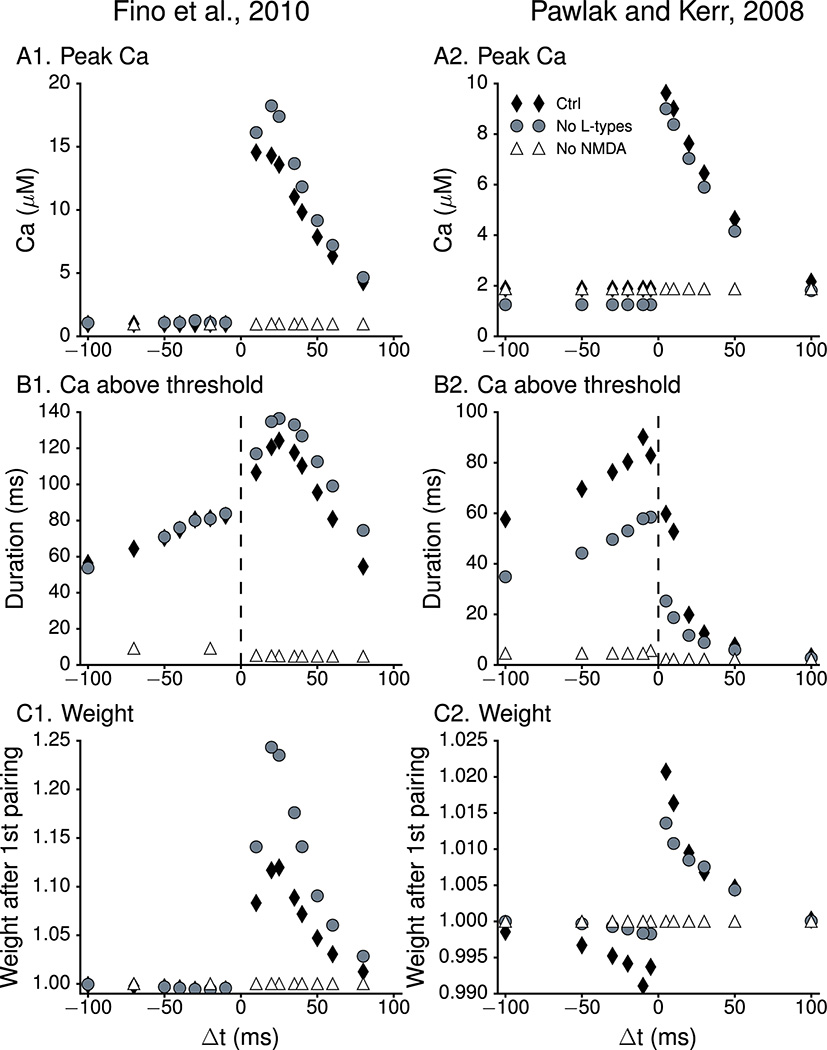
The striatum is a major site of learning and memory formation for sensorimotor and cognitive association. One of the mechanisms used by the brain for memory storage is synaptic plasticity - the long-lasting, activity-dependent change in synaptic strength. All forms of synaptic plasticity require an elevation in intracellular calcium, and a common hypothesis is that the amplitude and duration of calcium transients can determine the direction of synaptic plasticity. The utility of this hypothesis in the striatum is unclear in part because dopamine is required for striatal plasticity and in part because of the diversity in stimulation protocols. To test whether calcium can predict plasticity direction, we developed a calcium-based plasticity rule using a spiny projection neuron model with sophisticated calcium dynamics including calcium diffusion, buffering and pump extrusion. We utilized three spike timing-dependent plasticity (STDP) induction protocols, in which postsynaptic potentials are paired with precisely timed action potentials and the timing of such pairing determines whether potentiation or depression will occur. Results show that despite the variation in calcium dynamics, a single, calcium-based plasticity rule, which explicitly considers duration of calcium elevations, can explain the direction of synaptic weight change for all three STDP protocols. Additional simulations show that the plasticity rule correctly predicts the NMDA receptor dependence of long-term potentiation and the L-type channel dependence of long-term depression. By utilizing realistic calcium dynamics, the model reveals mechanisms controlling synaptic plasticity direction, and shows that the dynamics of calcium, not just calcium amplitude, are crucial for synaptic plasticity.
Keywords: basal ganglia; computational model; long-term potentiation/long-term depression; spike timing-dependent plasticity; striatum.
© 2016 Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




Similar articles
-
Interaction of inhibition and triplets of excitatory spikes modulates the NMDA-R-mediated synaptic plasticity in a computational model of spike timing-dependent plasticity.Hippocampus. 2013 Jan;23(1):75-86. doi: 10.1002/hipo.22057. Epub 2012 Jul 31. Hippocampus. 2013. PMID: 22851353
-
Corticostriatal paired-pulse potentiation produced by voltage-dependent activation of NMDA receptors and L-type Ca(2+) channels.J Neurophysiol. 2002 Jan;87(1):157-65. doi: 10.1152/jn.00115.2001. J Neurophysiol. 2002. PMID: 11784738
-
Spiking neurons, dopamine, and plasticity: timing is everything, but concentration also matters.Synapse. 2007 Jun;61(6):375-90. doi: 10.1002/syn.20378. Synapse. 2007. PMID: 17372980
-
Neurotransmitter roles in synaptic modulation, plasticity and learning in the dorsal striatum.Neuropharmacology. 2010 Jun;58(7):951-61. doi: 10.1016/j.neuropharm.2010.01.008. Epub 2010 Jan 21. Neuropharmacology. 2010. PMID: 20096294 Free PMC article. Review.
-
Spike-timing dependent plasticity in striatal interneurons.Neuropharmacology. 2011 Apr;60(5):780-8. doi: 10.1016/j.neuropharm.2011.01.023. Epub 2011 Jan 22. Neuropharmacology. 2011. PMID: 21262240 Review.
Cited by
-
Unraveling the dynamics of dopamine release and its actions on target cells.Trends Neurosci. 2023 Mar;46(3):228-239. doi: 10.1016/j.tins.2022.12.005. Epub 2023 Jan 10. Trends Neurosci. 2023. PMID: 36635111 Free PMC article. Review.
-
Basal ganglia mechanisms in action selection, plasticity, and dystonia.Eur J Paediatr Neurol. 2018 Mar;22(2):225-229. doi: 10.1016/j.ejpn.2018.01.005. Epub 2018 Jan 17. Eur J Paediatr Neurol. 2018. PMID: 29396175 Free PMC article. Review.
-
Thinking Outside the Box (and Arrow): Current Themes in Striatal Dysfunction in Movement Disorders.Neuroscientist. 2019 Aug;25(4):359-379. doi: 10.1177/1073858418807887. Epub 2018 Oct 31. Neuroscientist. 2019. PMID: 30379121 Free PMC article. Review.
-
Synaptic Plasticity Is Predicted by Spatiotemporal Firing Rate Patterns and Robust to In Vivo-like Variability.Biomolecules. 2022 Oct 1;12(10):1402. doi: 10.3390/biom12101402. Biomolecules. 2022. PMID: 36291612 Free PMC article.
-
Inhibition enhances spatially-specific calcium encoding of synaptic input patterns in a biologically constrained model.Elife. 2018 Oct 25;7:e38588. doi: 10.7554/eLife.38588. Elife. 2018. PMID: 30355449 Free PMC article.
References
-
- Allbritton NL, Meyer T, Stryer L. Range of messenger action of calcium ion and inositol 1,4,5-triphosphate. Science. 1992;258:1812–1815. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
