

Mitochondrial cAMP signaling
- PMID: 27233501
- PMCID: PMC5097110
- DOI: 10.1007/s00018-016-2282-2
Mitochondrial cAMP signaling
Abstract
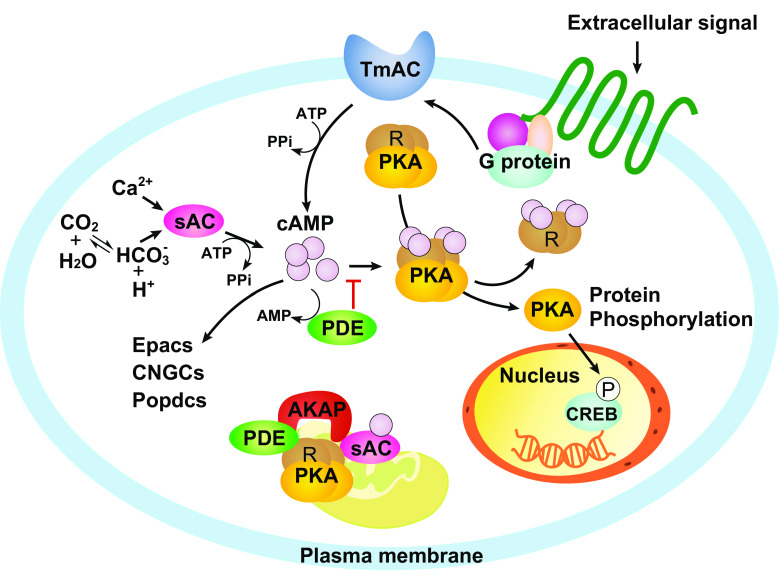
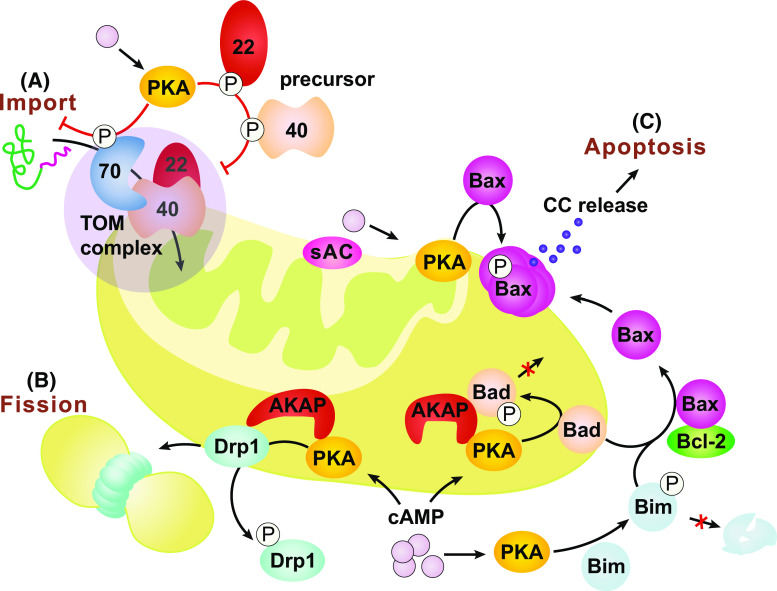
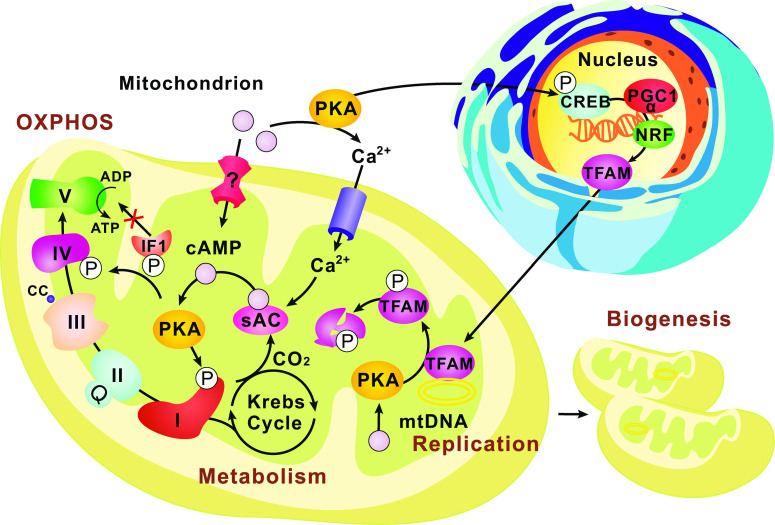
Cyclic adenosine 3, 5'-monophosphate (cAMP) is a ubiquitous second messenger regulating many biological processes, such as cell migration, differentiation, proliferation and apoptosis. cAMP signaling functions not only on the plasma membrane, but also in the nucleus and in organelles such as mitochondria. Mitochondrial cAMP signaling is an indispensable part of the cytoplasm-mitochondrion crosstalk that maintains mitochondrial homeostasis, regulates mitochondrial dynamics, and modulates cellular stress responses and other signaling pathways. Recently, the compartmentalization of mitochondrial cAMP signaling has attracted great attentions. This new input should be carefully taken into account when we interpret the findings of mitochondrial cAMP signaling. In this review, we summarize previous and recent progress in our understanding of mitochondrial cAMP signaling, including the components of the signaling cascade, and the function and regulation of this signaling pathway in different mitochondrial compartments.
Keywords: AKAP; BH3; Drp1; Fission; PKA; Protein import; TFAM; mtDNA.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
