

Chloroplast Membrane Remodeling during Freezing Stress Is Accompanied by Cytoplasmic Acidification Activating SENSITIVE TO FREEZING2
- PMID: 27233750
- PMCID: PMC4936559
- DOI: 10.1104/pp.16.00286
Chloroplast Membrane Remodeling during Freezing Stress Is Accompanied by Cytoplasmic Acidification Activating SENSITIVE TO FREEZING2
Abstract
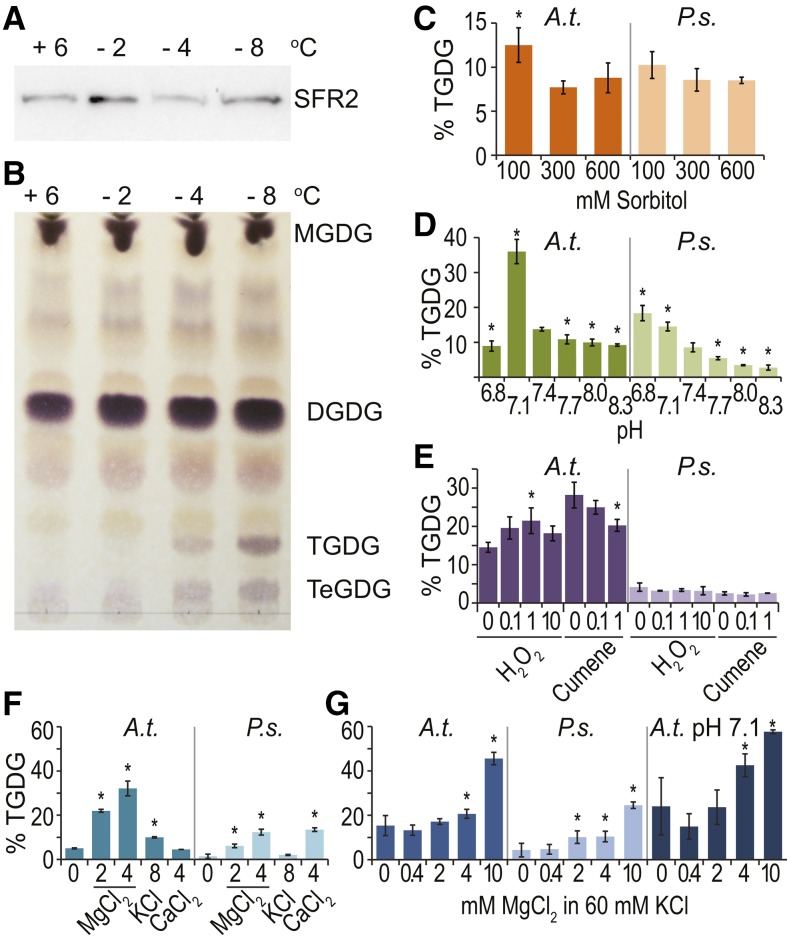
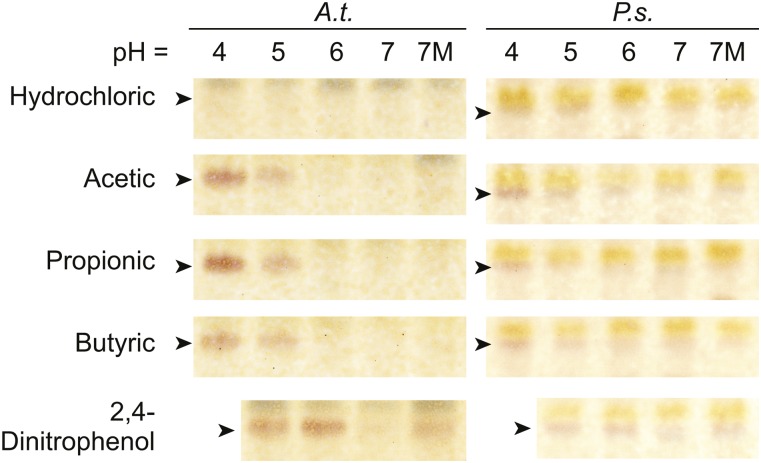
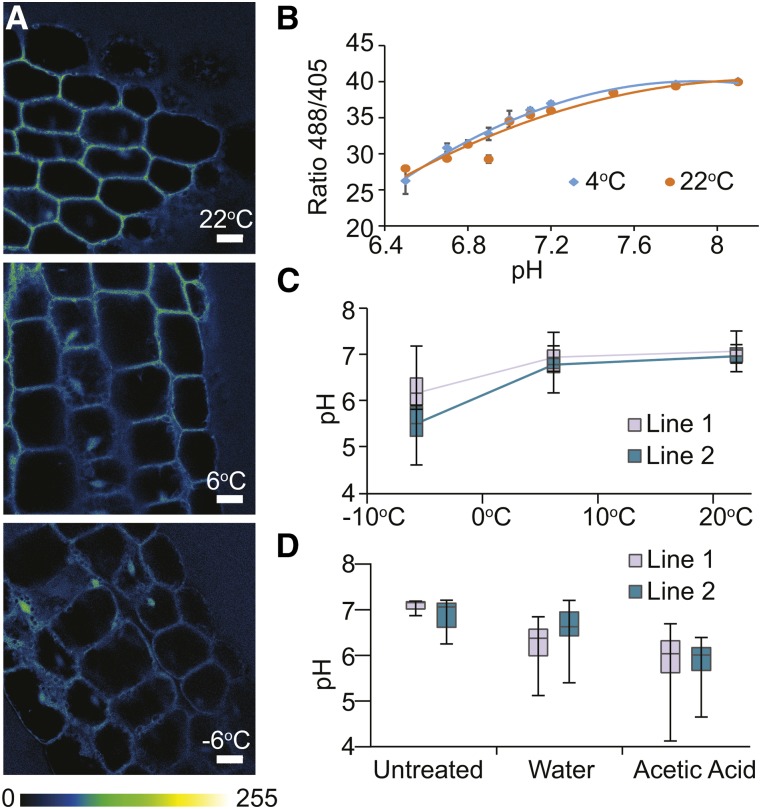
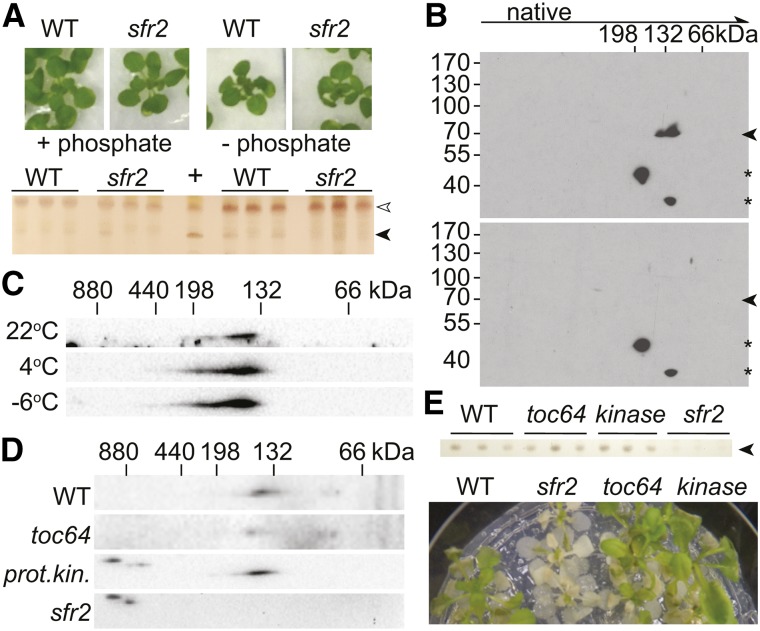
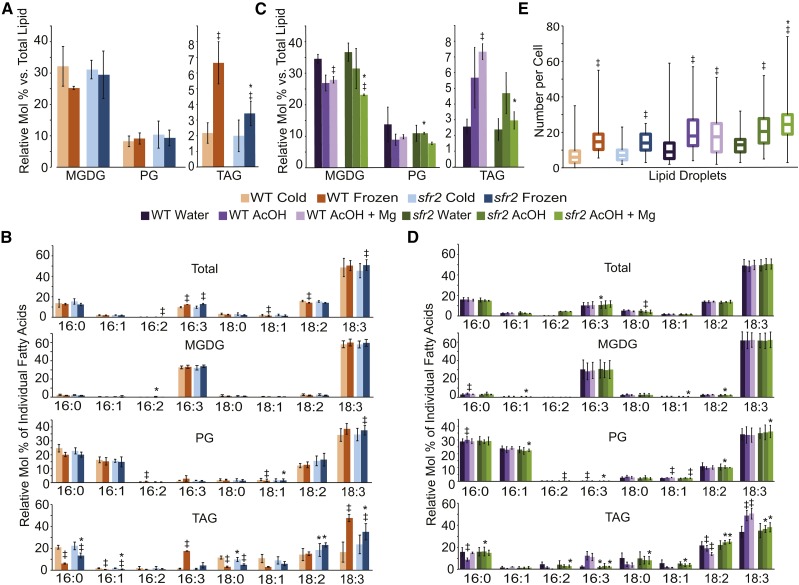
Low temperature is a seasonal abiotic stress that restricts native plant ranges and crop distributions. Two types of low-temperature stress can be distinguished: chilling and freezing. Much work has been done on the mechanisms by which chilling is sensed, but relatively little is known about how plants sense freezing. Recently, Arabidopsis (Arabidopsis thaliana) SENSITIVE TO FREEZING2 (SFR2) was identified as a protein that responds in a nontranscriptional manner to freezing. Here, we investigate the cellular conditions that allow SFR2 activation. Using a combination of isolated organelle, whole-tissue, and whole-plant assays, we provide evidence that SFR2 is activated by changes in cytosolic pH and Mg(2+) Manipulation of pH and Mg(2+) in cold-acclimated plants is shown to cause changes similar to those of freezing. We conclude that pH and Mg(2+) are perceived as intracellular cues as part of the sensing mechanism for freezing conditions. This evidence provides a specific molecular mechanism to combat freezing.
© 2016 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Aronsson H, Boij P, Patel R, Wardle A, Töpel M, Jarvis P (2007) Toc64/OEP64 is not essential for the efficient import of proteins into chloroplasts in Arabidopsis thaliana. Plant J 52: 53–68 - PubMed
-
- Browse J, Xin Z (2001) Temperature sensing and cold acclimation. Curr Opin Plant Biol 4: 241–246 - PubMed
-
- Bruce BD, Perry S, Froehlich J, Keegstra K (1994) In vitro import of protein into chloroplasts. In Gelvin SB, Schilperoort RA, eds, Plant Molecular Biology Manual. Kluwer Academic Publishers, Boston, pp 1–15
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
