

HSV-1 ICP27 targets the TBK1-activated STING signalsome to inhibit virus-induced type I IFN expression
- PMID: 27234299
- PMCID: PMC4931188
- DOI: 10.15252/embj.201593458
HSV-1 ICP27 targets the TBK1-activated STING signalsome to inhibit virus-induced type I IFN expression
Abstract
Herpes simplex virus (HSV) 1 stimulates type I IFN expression through the cGAS-STING-TBK1 signaling axis. Macrophages have recently been proposed to be an essential source of IFN during viral infection. However, it is not known how HSV-1 inhibits IFN expression in this cell type. Here, we show that HSV-1 inhibits type I IFN induction through the cGAS-STING-TBK1 pathway in human macrophages, in a manner dependent on the conserved herpesvirus protein ICP27. This viral protein was expressed de novo in macrophages with early nuclear localization followed by later translocation to the cytoplasm where ICP27 prevented activation of IRF3. ICP27 interacted with TBK1 and STING in a manner that was dependent on TBK1 activity and the RGG motif in ICP27. Thus, HSV-1 inhibits expression of type I IFN in human macrophages through ICP27-dependent targeting of the TBK1-activated STING signalsome.
Keywords: herpes simplex virus; immune evasion; innate immunity; type I IFN.
© 2016 The Authors.
Figures
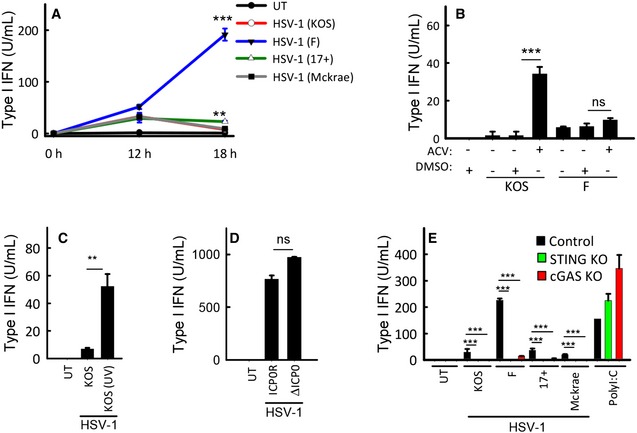
THP 1 cells were infected with the shown strains ofHSV ‐1 (MOI 3). Supernatants were harvested from untreated cultures or cells infected for 12 or 18 h for measurement of type IIFN bioactivity.THP 1 cells were treated with 0.1 μg/ml of acyclovir (ACV ) and infected with theKOS and FHSV ‐1 strains (MOI 3). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity.THP 1 cells were treated with infectious orUV ‐inactivatedHSV ‐1 (strainKOS ). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity.MDM s were infected withICP 0‐deficient or revertantHSV ‐1 (strainKOS ,MOI 3). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity.THP 1‐derived cells deficient forcGAS orSTING were infected with the shown strains ofHSV ‐1 (MOI 3) or stimulated with poly(I:C) (2 μg/ml). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity.
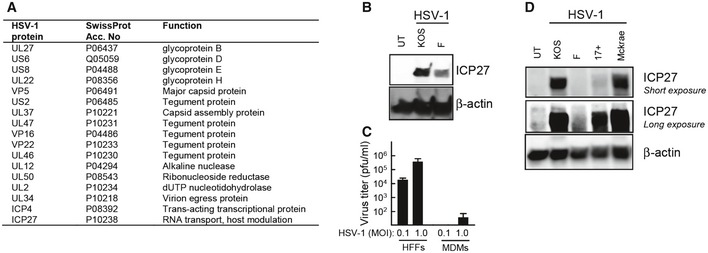
MDM s were infected for 18 h withHSV ‐1 (KOS ,MOI 1). Cytosolic viral proteins were detected by mass spectrometry usingiTRAQ labeling. Identified viral proteins are shown.MDM s were infected with theKOS or F strains ofHSV ‐1 (MOI 3). Cell lysates were harvested 12 hpi, and levels ofICP 27 and β‐actin were determined by Western blotting.HFF s andMDM s were infected withHSV ‐1 (KOS ,MOI 0.1, and 1.0). Culture medium was replaced 3 hpi and isolated 24 hpi for viral plaque assay. Data are presented as means of triplicates ±SD .THP 1 cells were infected with the indicated strains ofHSV ‐1 (MOI 3) for 12 h. Cell lysates were isolated, and levels ofICP 27 and β‐actin were determined by Western blotting.
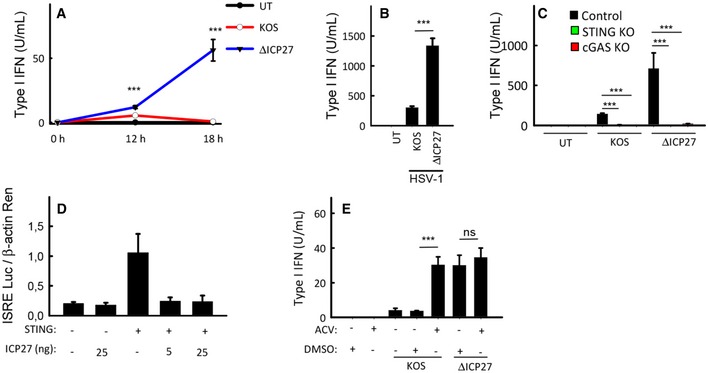
THP 1 cells were infected withHSV ‐1KOS or anICP 27‐deficient mutant (ΔICP 27) onKOS genetic background (MOI 3). Supernatants were harvested from untreated cultures or cells infected for 12 or 18 h for measurement of type IIFN bioactivity.MDM s were infected withHSV ‐1KOS or ΔICP 27 (MOI 3). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity.THP 1‐derived cells deficient forcGAS orSTING were infected withHSV ‐1KOS or ΔICP 27 (MOI 3). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity.HEK 293T cells were transfected with 12 ngSTING plasmidDNA , 5 or 25 ngICP 27 plasmidDNA , and reporter gene constructs as indicated (2 × 104 cells per well). Reporter gene activity was measured in lysates isolated 24 h post‐transfection.THP 1 cells were infected withHSV ‐1KOS or ΔICP 27 (MOI 3) in the presence or absence of acyclovir (ACV ). Supernatants were isolated 18 hpi for measurement of type IIFN bioactivity.
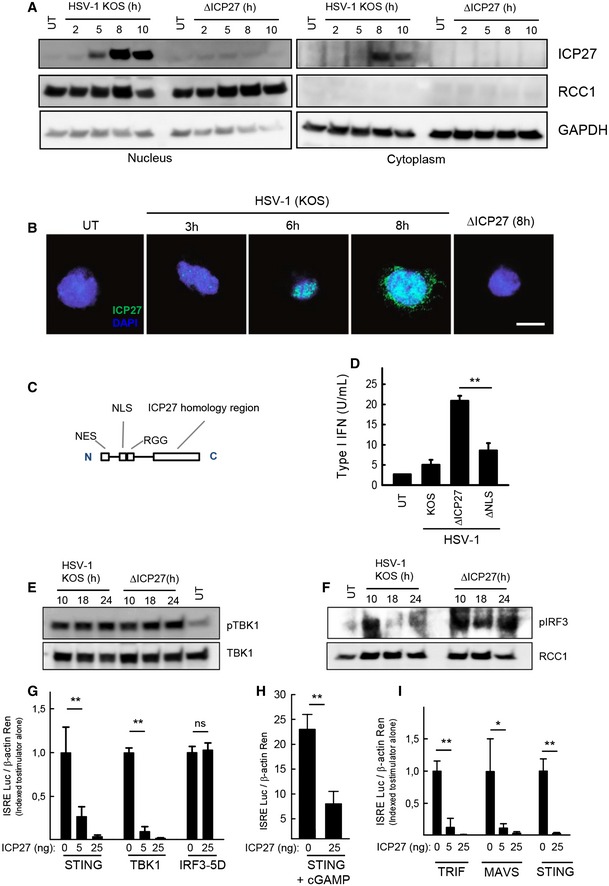
- A
THP 1 cells were infected withHSV ‐1KOS or ΔICP 27 (MOI 3). Cytoplasmic and nuclear extracts were isolated at the indicated time points post‐infection, and levels ofICP 27,RCC 1, andGAPDH were determined by Western blotting. - B
THP 1 cells were infected withHSV ‐1KOS or ΔICP 27 (MOI 10). The cells were fixed at the indicated time points post‐infection, stained withDAPI and an antibody againstICP 27, and visualized by confocal microscopy. Scale bar, 10 μm. - C, D
THP 1 cells were infected withHSV ‐1KOS , ΔICP 27, orHSV ‐1 ΔNLS . The localization of the majorNLS is illustrated in (C). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity. - E, F
THP 1 cells were infected withHSV ‐1KOS or ΔICP 27 (MOI 3). Cytoplasmic (E) and nuclear extracts (F) were isolated at the indicated time points post‐infection. Western blotting was used to determine levels of phospho‐ and totalTBK 1 were determined in the cytoplasmic fractions and levels of phospho‐IRF 3 andRCC 1 in the nuclear fractions. - G–I
HEK 293T cells were (G) transfected withSTING ,TBK 1,IRF 3‐5D, andICP 27, (H) transfected withSTING and stimulated for 16 h withcGAMP (4 μg/ml) or (I) transfected withTRIF ,MAVS ,STING , andICP 27, together with the reporter gene constructs indicated. Reporter gene activity was measured 24 h post‐transfection.
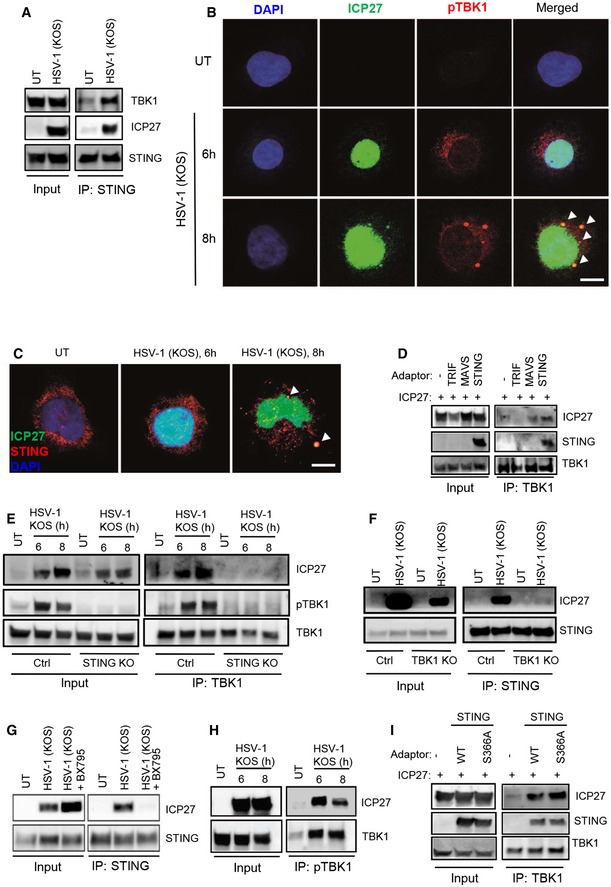
- A
Lysates from
THP 1 cells infected withHSV ‐1KOS (MOI 10) were subjected to immunoprecipitation using an anti‐STING antibody, and the presence ofTBK 1,STING , andICP 27 in the precipitates was detected by Western blotting. - B, C
THP 1 cells were infected withHSV ‐1KOS (MOI 10) for 6 and 8 h. The cells were fixed and stained withDAPI and anti‐ICP 27 together with anti‐pTBK 1 (B), or anti‐STING (C). The stainings were visualized by confocal microscopy. Arrowheads, co‐localizations betweenICP 27 andpTBK 1/STING . Scale bar, 10 μm. - D
TBK 1 was immunoprecipitated from whole‐cell lysates fromHEK 293T cells transfected withICP 27 and each of the adaptor proteinsTRIF ,MAVS , andSTING . The precipitates were immunoblotted forTBK 1,ICP 27, andSTING . - E
THP 1 control andSTING KO cells were infected withHSV ‐1KOS (MOI 10) for 6 and 8 h, and total lysates were generated.TBK 1 was immunoprecipitated and subjected to Western blotting using antibodies againstICP 27, phospho‐TBK 1 (pTBK 1), andTBK 1. - F
THP 1 control andTBK 1KO cells were infected withHSV ‐1KOS (MOI 10) for 8 h, and total lysates were generated.STING was immunoprecipitated and subjected to Western blotting using antibodies againstICP 27 andSTING . - G
THP 1 cells were infected withHSV ‐1KOS (MOI 10) for 8 h in the presence or absence of theTBK 1 inhibitorBX 795 (BX , 200 nM). Total lysates were generated andSTING was immunoprecipitated and subjected to Western blotting using antibodies againstICP 27 andSTING . - H
THP 1 cells were infected withHSV ‐1KOS (MOI 10) for 6 and 8 h, and total lysates were generated. Phospho‐TBK 1 was immunoprecipitated and subjected to Western blotting using antibodies againstICP 27 and totalTBK 1. - I
TBK 1 was immunoprecipitated from whole‐cell lysates fromHEK 293T cells transfected withICP 27 andSTING (eitherWT or S366A). The precipitates were immunoblotted forTBK 1,ICP 27, and STING.
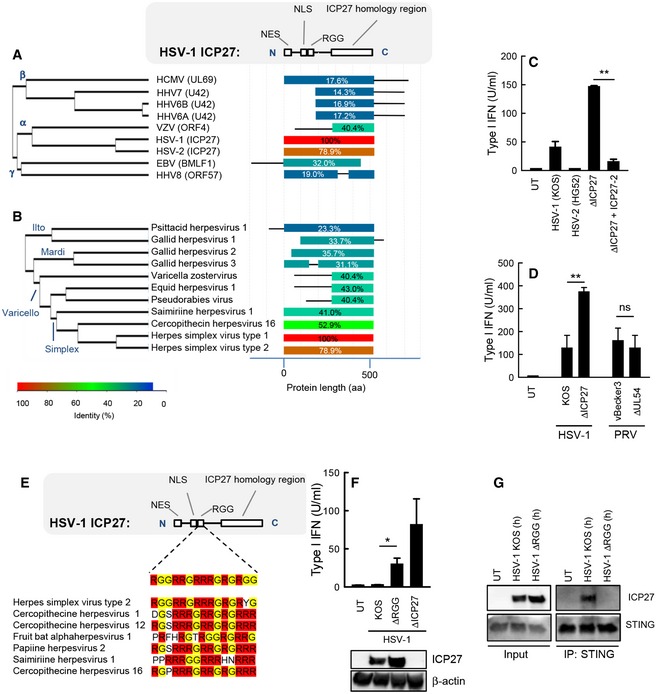
- A, B
Alignment of the
ICP 27 homologs of the human herpesviruses (A) and a series of alpha‐herpesviruses (B). The diagram to the right illustrates the degree of similarity withHSV ‐1ICP 27 and the distribution within the molecules. - C
THP 1 cells were infected withHSV ‐1 (KOS ),HSV ‐2 (HG 52),HSV ‐1 ΔICP 27, orHSV ‐1 ΔICP 27 rescued withHSV ‐2ICP 27 (MOI 3). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity. - D
RAW 264.7 cells were infected withHSV ‐1 (KOS ),PRV (vB ecker3) and the corresponding mutants lackingICP 27 (UL 54 in the case ofPRV ). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity. - E
Alignment of the
RGG box of inICP 27 proteins of the simplex virus genera of alpha‐herpesviruses. - F
THP 1 cells were infected withHSV ‐1 (KOS ), ΔRGG , or ΔICP 27 (MOI 3). Supernatants were harvested 18 hpi for measurement of type IIFN bioactivity.ICP 27 expression in the infected cells was determined by Western blotting. - G
THP 1 cells were infected withHSV ‐1KOS orHSV ‐1 ΔRGG (MOI 10) for 8 h, and total lysates were generated.STING was immunoprecipitated and subjected to Western blotting using antibodies againstICP 27 andSTING .
References
-
- Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124: 783–801 - PubMed
-
- Andersen LL, Mork N, Reinert LS, Kofod‐Olsen E, Narita R, Jorgensen SE, Skipper KA, Honing K, Gad HH, Ostergaard L, Orntoft TF, Hornung V, Paludan SR, Mikkelsen JG, Fujita T, Christiansen M, Hartmann R, Mogensen TH (2015) Functional IRF3 deficiency in a patient with herpes simplex encephalitis. J Exp Med 212: 1371–1379 - PMC - PubMed
-
- Chaurushiya MS, Lilley CE, Aslanian A, Meisenhelder J, Scott DC, Landry S, Ticau S, Boutell C, Yates JR III, Schulman BA, Hunter T, Weitzman MD (2012) Viral E3 ubiquitin ligase‐mediated degradation of a cellular E3: viral mimicry of a cellular phosphorylation mark targets the RNF8 FHA domain. Mol Cell 46: 79–90 - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
