

Pre-mRNA splicing is modulated by antifungal drugs in the filamentous fungus Neurospora crassa
- PMID: 27239448
- PMCID: PMC4821360
- DOI: 10.1002/2211-5463.12047
Pre-mRNA splicing is modulated by antifungal drugs in the filamentous fungus Neurospora crassa
Abstract
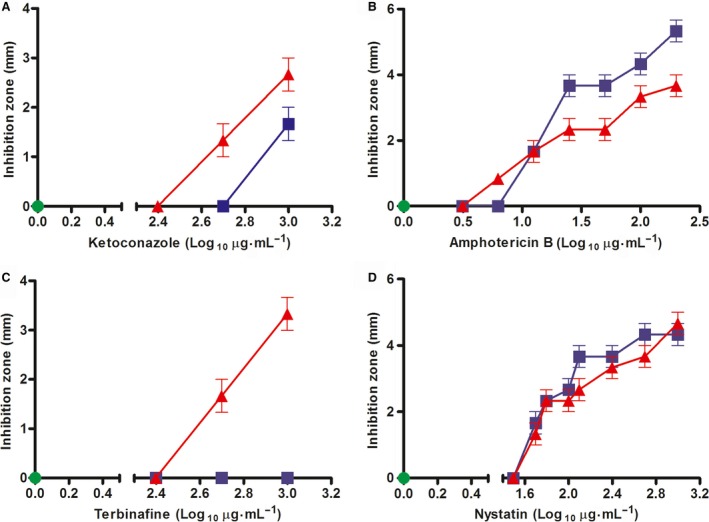
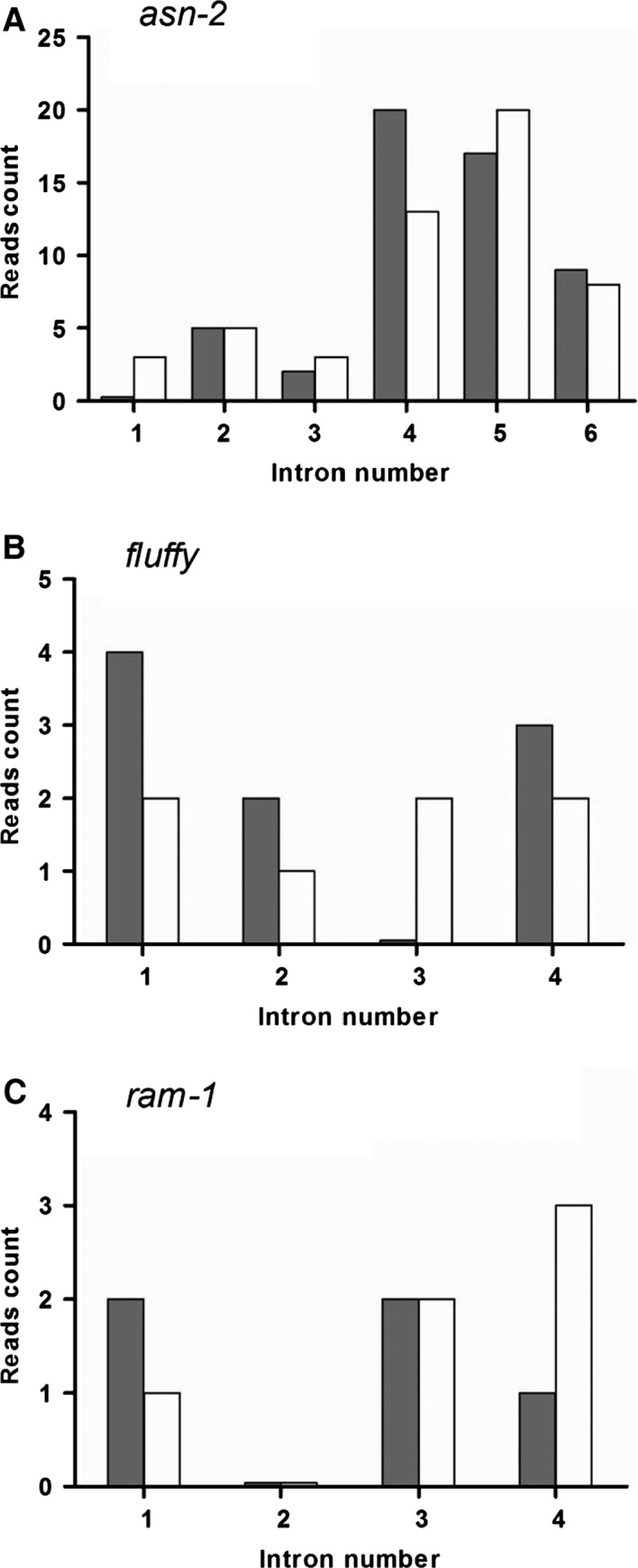
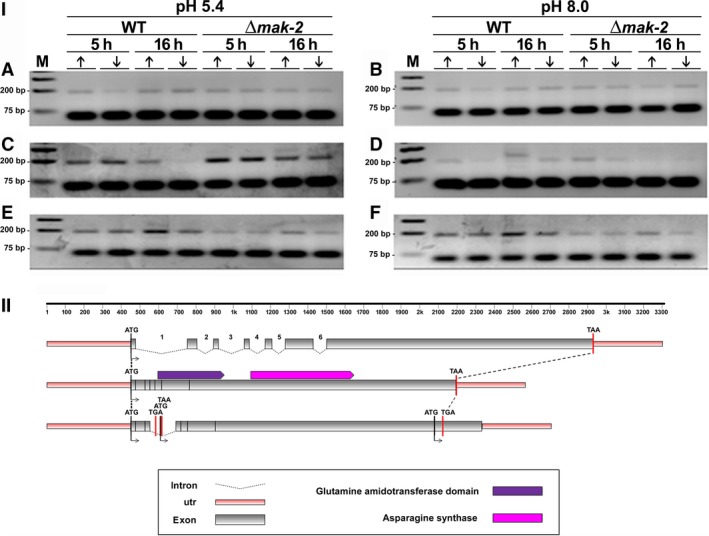
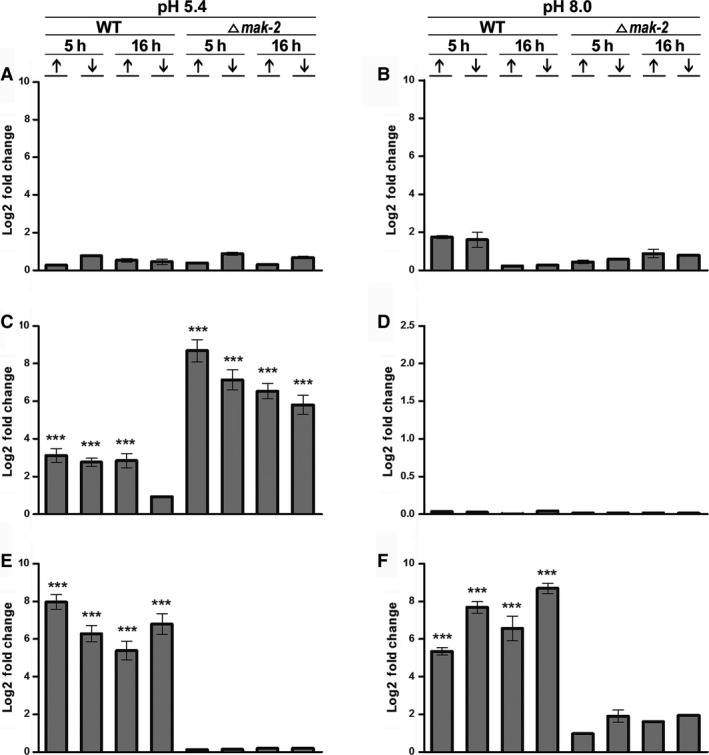
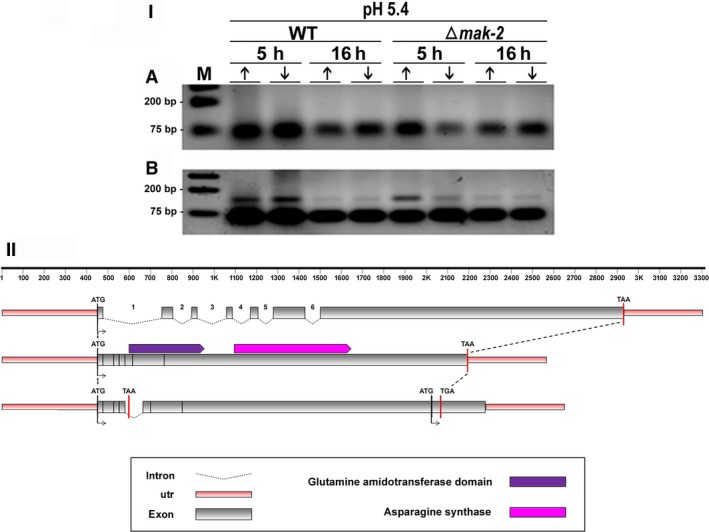
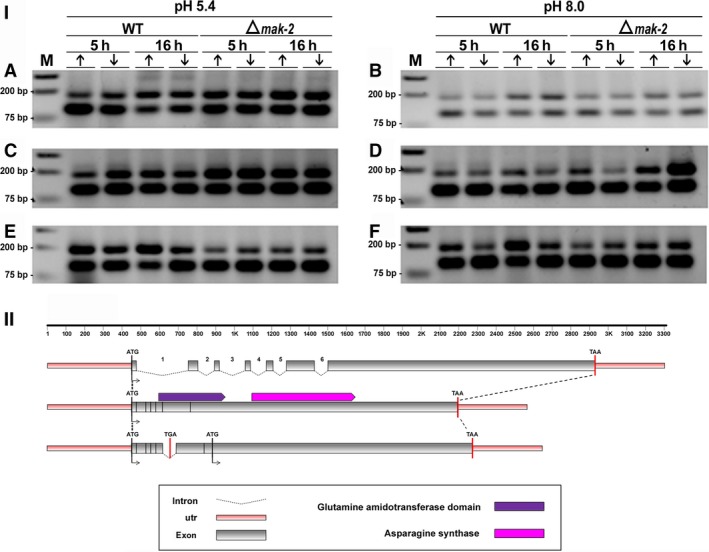
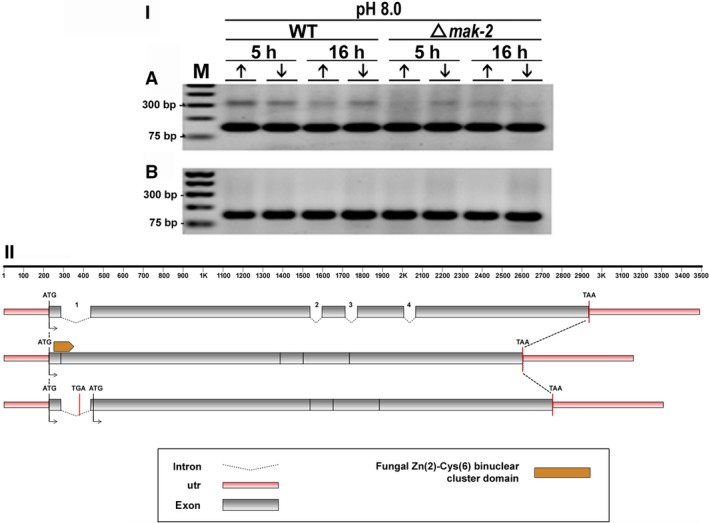
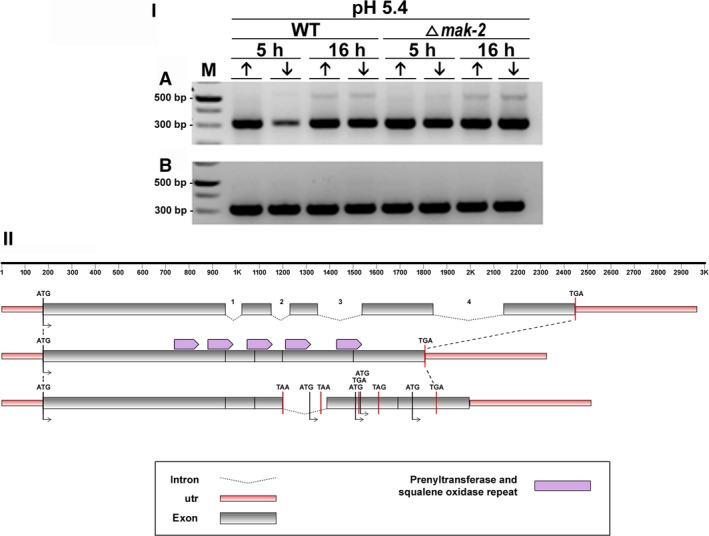
For this study, we sought to identify pre-mRNA processing events modulated by changes in extracellular pH, inorganic phosphate, and antifungal drugs. We examined genes with at least four putative introns whose transcriptional level responded to these effectors. We showed that the intron retention levels of genes encoding asparagine synthetase 2, C6-zinc finger regulator (fluffy), and a farnesyltransferase respond to amphotericin B, ketoconazole, and other effectors. In general, the assayed antifungals promoted the disruption of the structural domains of these proteins probably leading to their inactivation, which emphasize the complexity of the metabolic modulation exerted by antifungal signaling.
Keywords: Neurospora crassa; Pi regulation; alternative splicing; amphotericin B; asparagine synthetase; intron retention; ketoconazole.
Figures
References
-
- Kempken F (2013) Alternative splicing in ascomycetes. Appl Microbiol Biotechnol 97, 4235–4241. - PubMed
-
- Strandberg R, Tzelepis G, Johannesson H and Karlsson M (2013) Coexistence and expression profiles of two alternative splice variants of the pheromone receptor gene pre1 in Neurospora crassa . Arch Microbiol 195, 773–780. - PubMed
-
- Geng C and Paukstelis PJ (2014) An in vitro peptide complementation assay for CYT‐18‐dependent group I intron splicing reveals a new role for the N‐terminus. Biochemistry 53, 1311–1319. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
