

Evolution and diversity of floral scent chemistry in the euglossine bee-pollinated orchid genus Gongora
- PMID: 27240855
- PMCID: PMC4934395
- DOI: 10.1093/aob/mcw072
Evolution and diversity of floral scent chemistry in the euglossine bee-pollinated orchid genus Gongora
Abstract
•
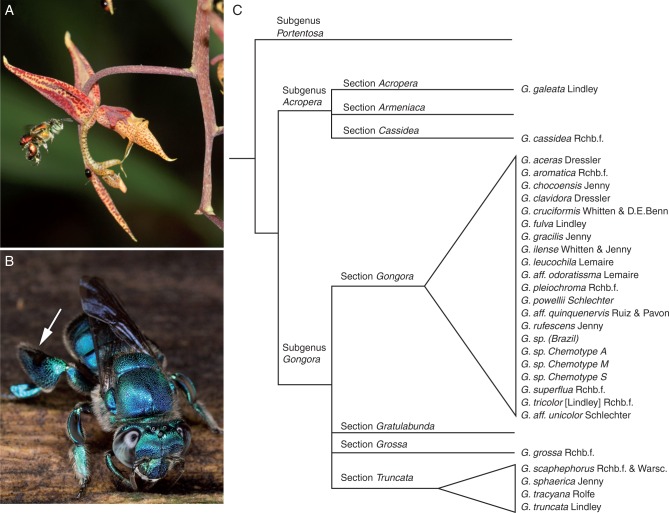
Background and aims: Animal-pollinated angiosperms have evolved a variety of signalling mechanisms to attract pollinators. Floral scent is a key component of pollinator attraction, and its chemistry modulates both pollinator behaviour and the formation of plant-pollinator networks. The neotropical orchid genus Gongora exhibits specialized pollinator associations with male orchid bees (Euglossini). Male bees visit orchid flowers to collect volatile chemical compounds that they store in hind-leg pouches to use subsequently during courtship display. Hence, Gongora floral scent compounds simultaneously serve as signalling molecules and pollinator rewards. Furthermore, because floral scent acts as the predominant reproductive isolating barrier among lineages, it has been hypothesized that chemical traits are highly species specific. A comparative analysis of intra- and inter-specific variation of floral scent chemistry was conducted to investigate the evolutionary patterns across the genus. •
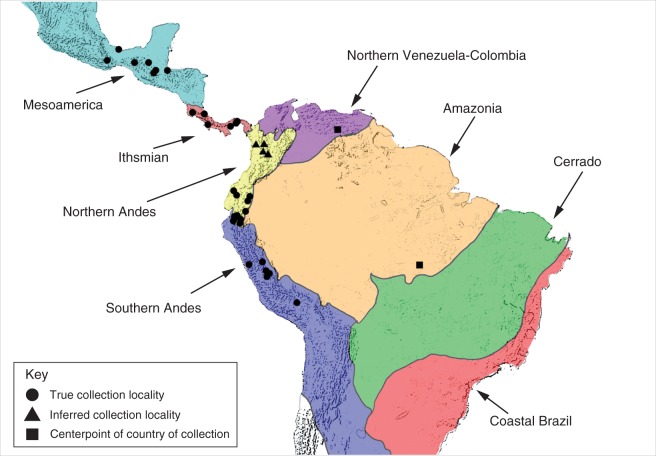
Methods: Gas chromatography-mass spectrometry (GC-MS) was used to analyse the floral scent of 78 individuals belonging to 28 different species of Gongora from two of the three major lineages sampled across the neotropical region. Multidimensional scaling and indicator value analyses were implemented to investigate the patterns of chemical diversity within and among taxonomic groups at various geographic scales. Additionally, pollinator observations were conducted on a sympatric community of Gongora orchids exhibiting distinct floral scent phenotypes. •
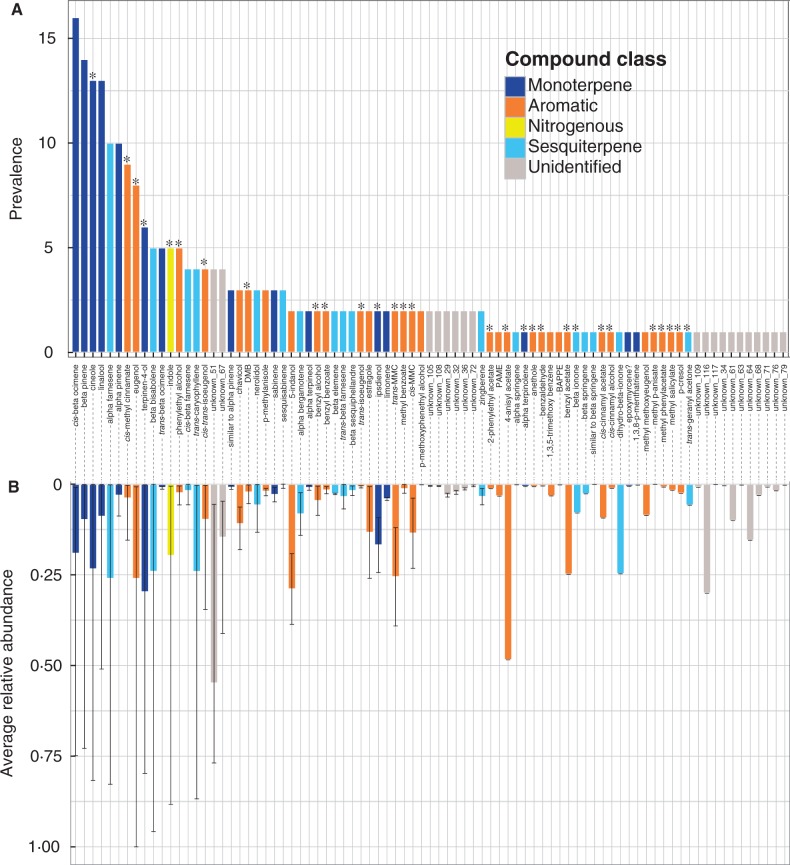
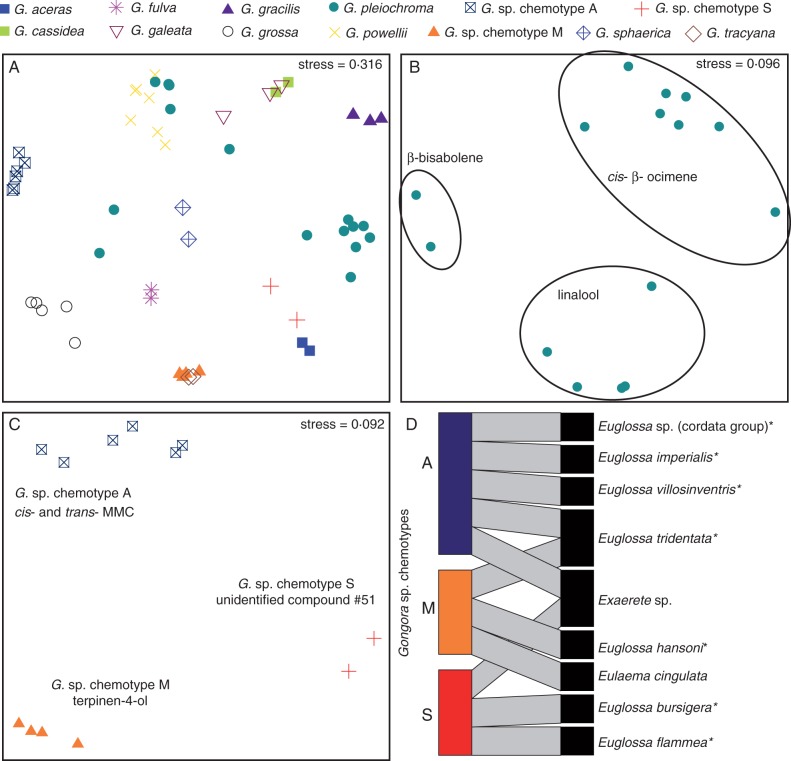
Key results: A total of 83 floral volatiles, mainly terpenes and aromatic compounds, were detected. Many of the identified compounds are common across diverse angiosperm families (e.g. cineole, eugenol, β-ocimene, β-pinene and terpinen-4-ol), while others are relatively rare outside euglossine bee-pollinated orchid lineages. Additionally, 29 volatiles were identified that are known to attract and elicit collection behaviour in male bees. Floral scent traits were less variable within species than between species, and the analysis revealed exceptional levels of cryptic diversity. Gongora species were divided into 15 fragrance groups based on shared compounds. Fragrance groups indicate that floral scent variation is not predicted by taxonomic rank or biogeographic region. •
Conclusions: Gongora orchids emit a diverse array of scent molecules that are largely species specific, and closely related taxa exhibit qualitatively and quantitatively divergent chemical profiles. It is shown that within a community, Gongora scent chemotypes are correlated with near non-overlapping bee pollinator assemblies. The results lend support to the hypothesis that floral scent traits regulate the architecture of bee pollinator associations. Thus, Gongora provides unique opportunities to examine the interplay between floral traits and pollinator specialization in plant-pollinator mutualisms.
Keywords: Euglossa; Euglossine bees; floral scent; orchid genus Gongora; plant–pollinator mutualism.
© The Author 2016. Published by Oxford University Press on behalf of the Annals of Botany Company.
Figures
Comment in
-
Orchid conservation: further links.Ann Bot. 2016 Jul;118(1):89-91. doi: 10.1093/aob/mcw147. Ann Bot. 2016. PMID: 27385771 Free PMC article.
References
-
- Ackerman JD. 1989. Geographic and seasonal variation in fragrance choices and preferences of male euglossine bees. Biotropica 21: 340–347.
-
- Adams RP. 2007. Identification of essential oil components by gas chromatography/mass spectrometry. Carol Stream, IL: Allured Publishing Corporation.
-
- Allen P. 1954. Pollinatioin in Gongora maculata. Ceiba 4: 121–125.
-
- Beals EW. 1984. Bray–Curtis ordination – an effective strategy for analysis of multivariate ecological data. Advances in Ecological Research 14: 1–55.
-
- Breitkopf H, Onstein RE, Cafasso D, Schlueter PM, Cozzolino S. 2015. Multiple shifts to different pollinators fuelled rapid diversification in sexually deceptive Ophrys orchids. New Phytologist 207: 377–389. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
