

Protective efficacy and safety of liver stage attenuated malaria parasites
- PMID: 27241521
- PMCID: PMC4886212
- DOI: 10.1038/srep26824
Protective efficacy and safety of liver stage attenuated malaria parasites
Abstract
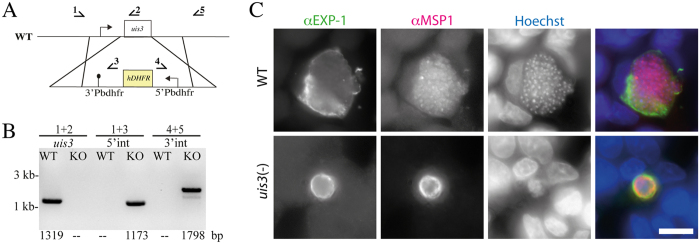
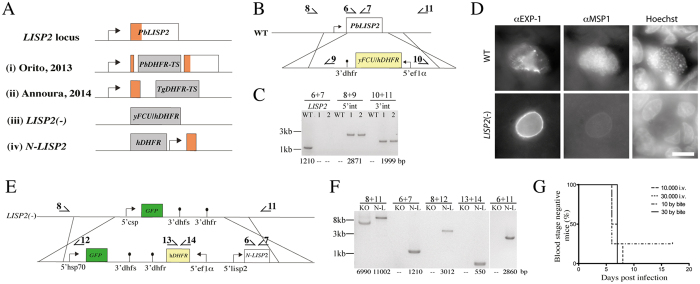
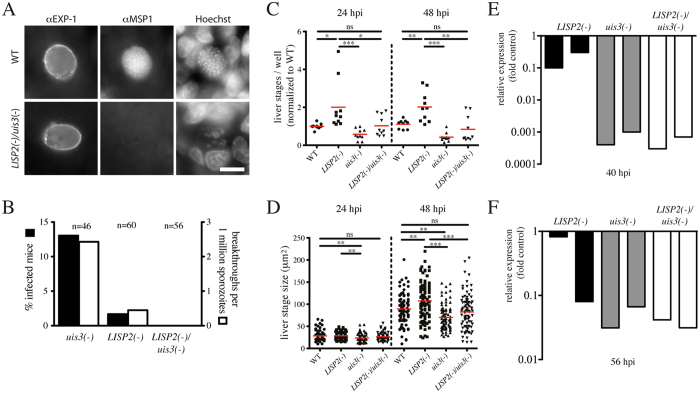
During the clinically silent liver stage of a Plasmodium infection the parasite replicates from a single sporozoite into thousands of merozoites. Infection of humans and rodents with large numbers of sporozoites that arrest their development within the liver can cause sterile protection from subsequent infections. Disruption of genes essential for liver stage development of rodent malaria parasites has yielded a number of attenuated parasite strains. A key question to this end is how increased attenuation relates to vaccine efficacy. Here, we generated rodent malaria parasite lines that arrest during liver stage development and probed the impact of multiple gene deletions on attenuation and protective efficacy. In contrast to P. berghei strain ANKA LISP2(-) or uis3(-) single knockout parasites, which occasionally caused breakthrough infections, the double mutant lacking both genes was completely attenuated even when high numbers of sporozoites were administered. However, different vaccination protocols showed that LISP2(-) parasites protected better than uis3(-) and double mutants. Hence, deletion of several genes can yield increased safety but might come at the cost of protective efficacy.
Figures
Similar articles
-
Engineering of Genetically Arrested Parasites (GAPs) For a Precision Malaria Vaccine.Front Cell Infect Microbiol. 2017 May 31;7:198. doi: 10.3389/fcimb.2017.00198. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28620583 Free PMC article. Review.
-
Genetically modified Plasmodium parasites as a protective experimental malaria vaccine.Nature. 2005 Jan 13;433(7022):164-7. doi: 10.1038/nature03188. Epub 2004 Dec 5. Nature. 2005. PMID: 15580261
-
A new approach to generate a safe double-attenuated Plasmodium liver stage vaccine.Int J Parasitol. 2013 May;43(6):503-14. doi: 10.1016/j.ijpara.2013.01.010. Epub 2013 Mar 13. Int J Parasitol. 2013. PMID: 23500072
-
Comparative efficacy of pre-erythrocytic whole organism vaccine strategies against the malaria parasite.Vaccine. 2011 Sep 16;29(40):7002-8. doi: 10.1016/j.vaccine.2011.07.034. Epub 2011 Jul 23. Vaccine. 2011. PMID: 21787828
-
Genetically modified Plasmodium highlights the potential of whole parasite vaccine strategies.Trends Immunol. 2005 Jun;26(6):295-7. doi: 10.1016/j.it.2005.04.005. Trends Immunol. 2005. PMID: 15922944 Review.
Cited by
-
A Plasmodium Parasite with Complete Late Liver Stage Arrest Protects against Preerythrocytic and Erythrocytic Stage Infection in Mice.Infect Immun. 2018 Apr 23;86(5):e00088-18. doi: 10.1128/IAI.00088-18. Print 2018 May. Infect Immun. 2018. PMID: 29440367 Free PMC article.
-
A Plasmodium late liver stage arresting GAP provides superior protection in mice.NPJ Vaccines. 2024 Oct 18;9(1):193. doi: 10.1038/s41541-024-00975-0. NPJ Vaccines. 2024. PMID: 39424860 Free PMC article.
-
The s48/45 six-cysteine proteins: mediators of interaction throughout the Plasmodium life cycle.Int J Parasitol. 2017 Jun;47(7):409-423. doi: 10.1016/j.ijpara.2016.10.002. Epub 2016 Nov 27. Int J Parasitol. 2017. PMID: 27899328 Free PMC article. Review.
-
Transport mechanisms at the malaria parasite-host cell interface.PLoS Pathog. 2021 Apr 1;17(4):e1009394. doi: 10.1371/journal.ppat.1009394. eCollection 2021 Apr. PLoS Pathog. 2021. PMID: 33793667 Free PMC article. Review.
-
Engineering of Genetically Arrested Parasites (GAPs) For a Precision Malaria Vaccine.Front Cell Infect Microbiol. 2017 May 31;7:198. doi: 10.3389/fcimb.2017.00198. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28620583 Free PMC article. Review.
References
-
- Nussenzweig R. S., Vanderberg J., Most H. & Orton C. Protective immunity produced by the injection of x-irradiated sporozoites of Plasmodium berghei. Nature 216, 160–162 (1967). - PubMed
-
- Silvie O. et al.. Effects of irradiation on Plasmodium falciparum sporozoite hepatic development: implications for the design of pre-erythrocytic malaria vaccines. Parasite Immunol. 24, 221–223 (2002). - PubMed
-
- Seder R. A. et al.. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 341, 1359–1365 (2013). - PubMed
-
- Hoffman S. L. et al.. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum sporozoites. J. Infect. Dis. 185, 1155–1164 (2002). - PubMed
-
- Beaudouin R. L., Strome C. P. A., Mitchell F. & Tubergen T. A. Plasmodium berghei: Immunization of mice against the ANKA strain using the unaltered sporozoite as an antigen. Exp. Parasitol. 42, 1–5 (1977). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
