

MICOS and phospholipid transfer by Ups2-Mdm35 organize membrane lipid synthesis in mitochondria
- PMID: 27241913
- PMCID: PMC4896057
- DOI: 10.1083/jcb.201602007
MICOS and phospholipid transfer by Ups2-Mdm35 organize membrane lipid synthesis in mitochondria
Abstract
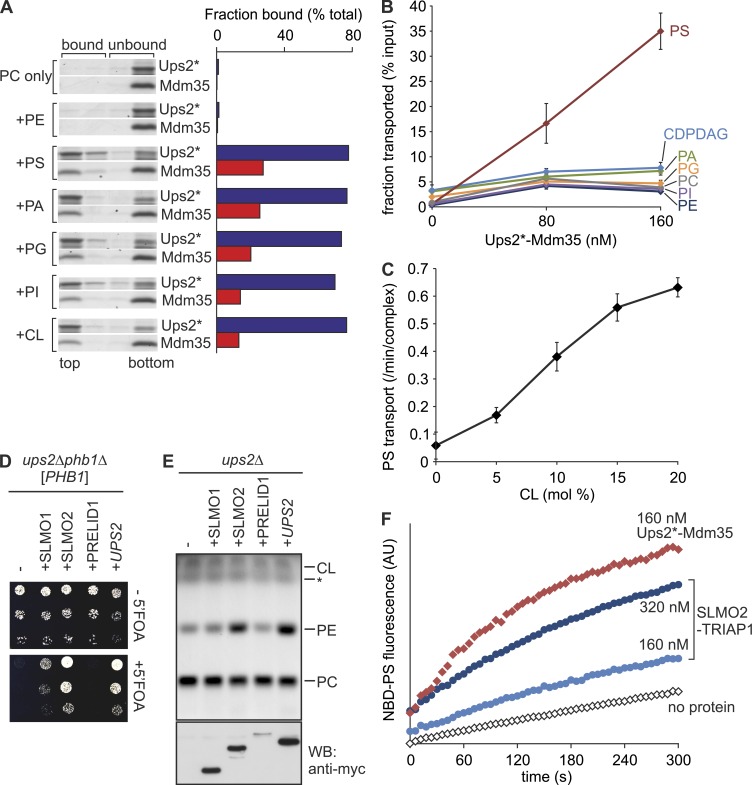
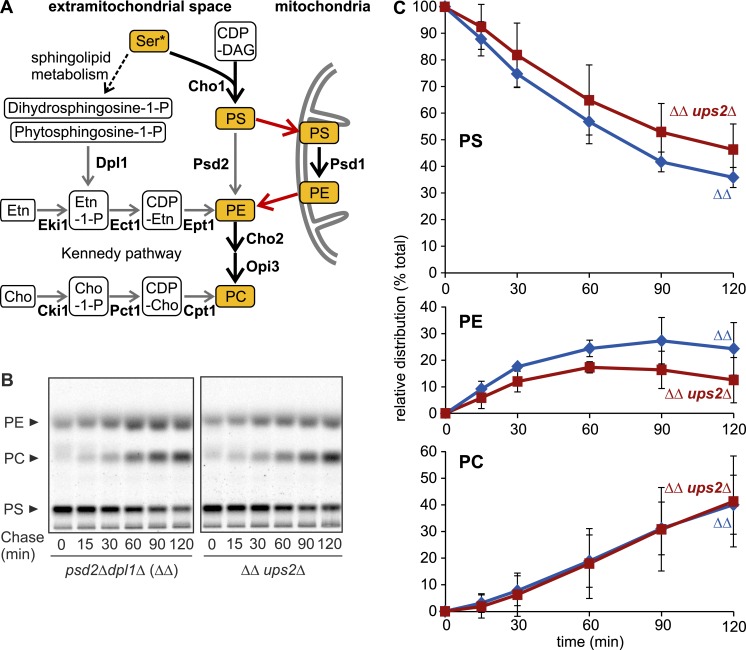
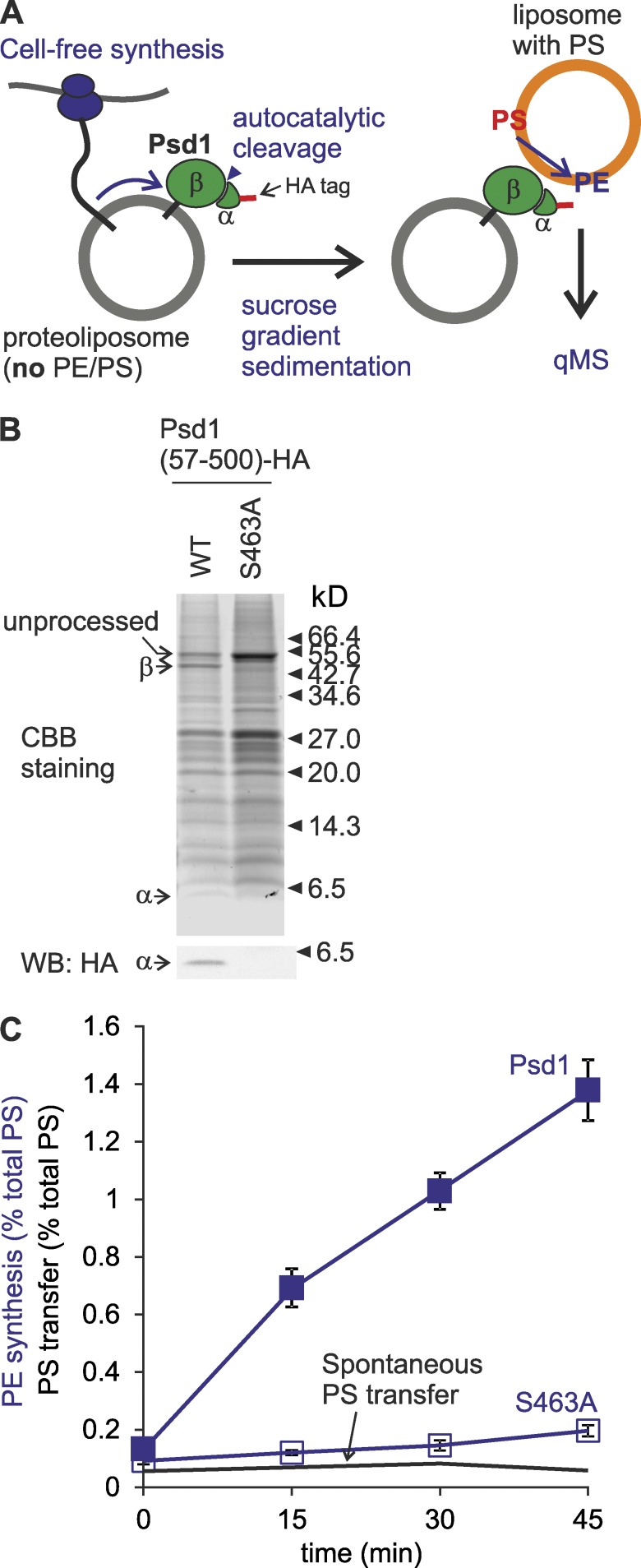
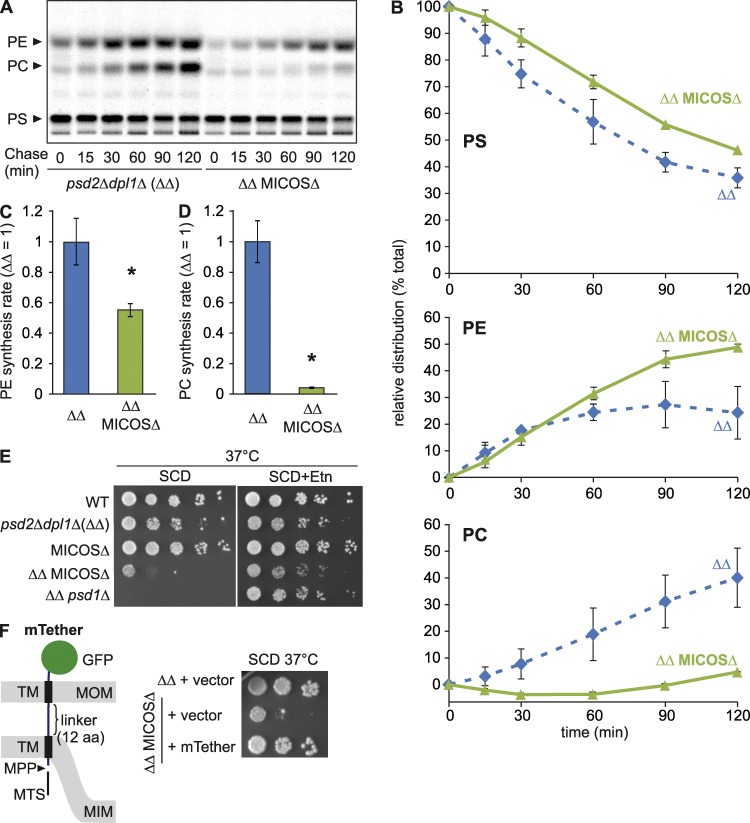
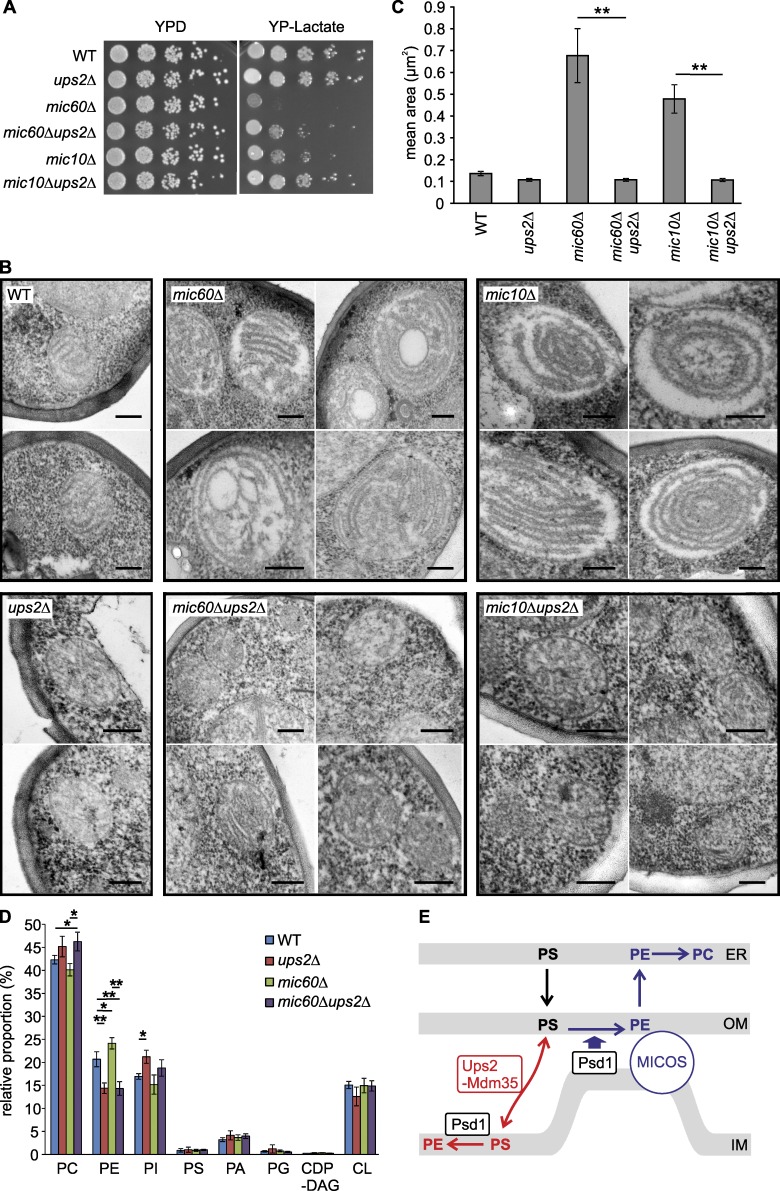
Mitochondria exert critical functions in cellular lipid metabolism and promote the synthesis of major constituents of cellular membranes, such as phosphatidylethanolamine (PE) and phosphatidylcholine. Here, we demonstrate that the phosphatidylserine decarboxylase Psd1, located in the inner mitochondrial membrane, promotes mitochondrial PE synthesis via two pathways. First, Ups2-Mdm35 complexes (SLMO2-TRIAP1 in humans) serve as phosphatidylserine (PS)-specific lipid transfer proteins in the mitochondrial intermembrane space, allowing formation of PE by Psd1 in the inner membrane. Second, Psd1 decarboxylates PS in the outer membrane in trans, independently of PS transfer by Ups2-Mdm35. This latter pathway requires close apposition between both mitochondrial membranes and the mitochondrial contact site and cristae organizing system (MICOS). In MICOS-deficient cells, limiting PS transfer by Ups2-Mdm35 and reducing mitochondrial PE accumulation preserves mitochondrial respiration and cristae formation. These results link mitochondrial PE metabolism to MICOS, combining functions in protein and lipid homeostasis to preserve mitochondrial structure and function.
© 2016 Aaltonen et al.
Figures
References
-
- Alkhaja A.K., Jans D.C., Nikolov M., Vukotic M., Lytovchenko O., Ludewig F., Schliebs W., Riedel D., Urlaub H., Jakobs S., and Deckers M.. 2012. MINOS1 is a conserved component of mitofilin complexes and required for mitochondrial function and cristae organization. Mol. Biol. Cell. 23:247–257. 10.1091/mbc.E11-09-0774 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
