

Redox biology and the interface between bioenergetics, autophagy and circadian control of metabolism
- PMID: 27242268
- PMCID: PMC5124549
- DOI: 10.1016/j.freeradbiomed.2016.05.022
Redox biology and the interface between bioenergetics, autophagy and circadian control of metabolism
Abstract
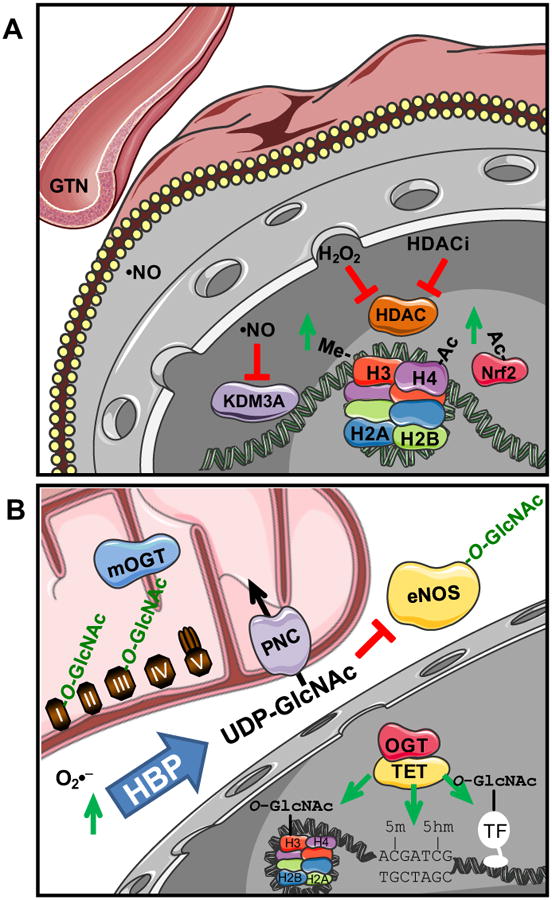
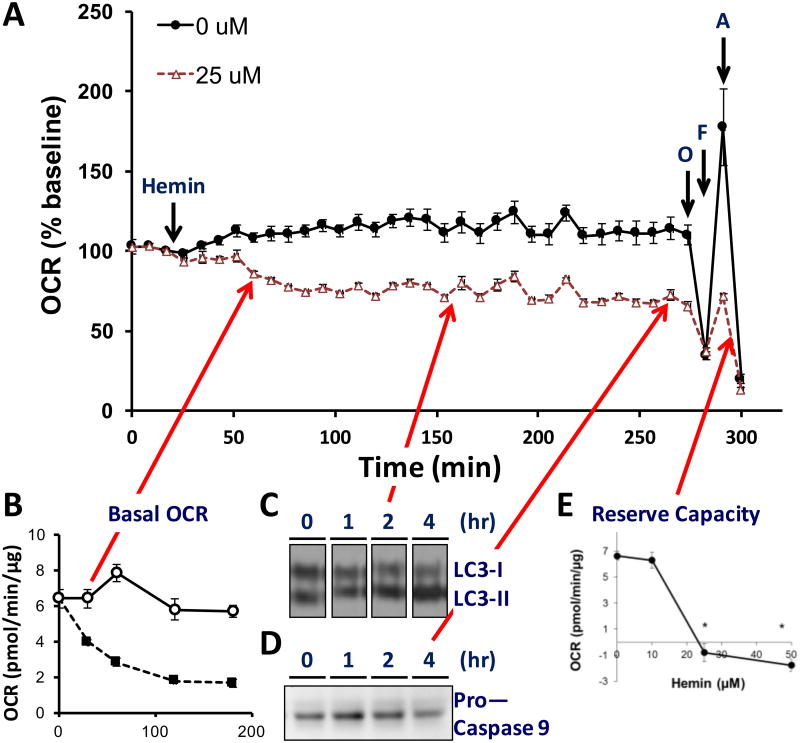
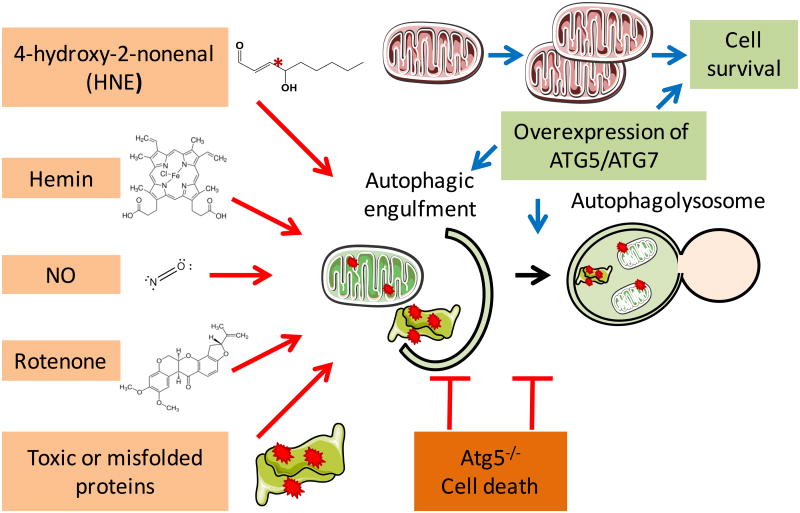
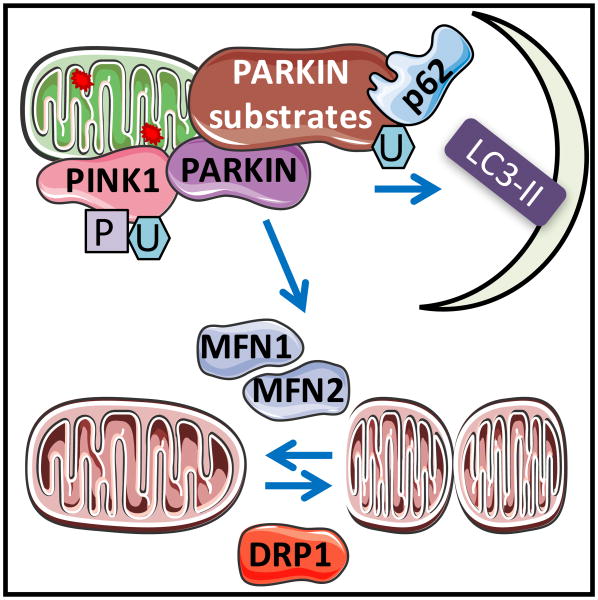
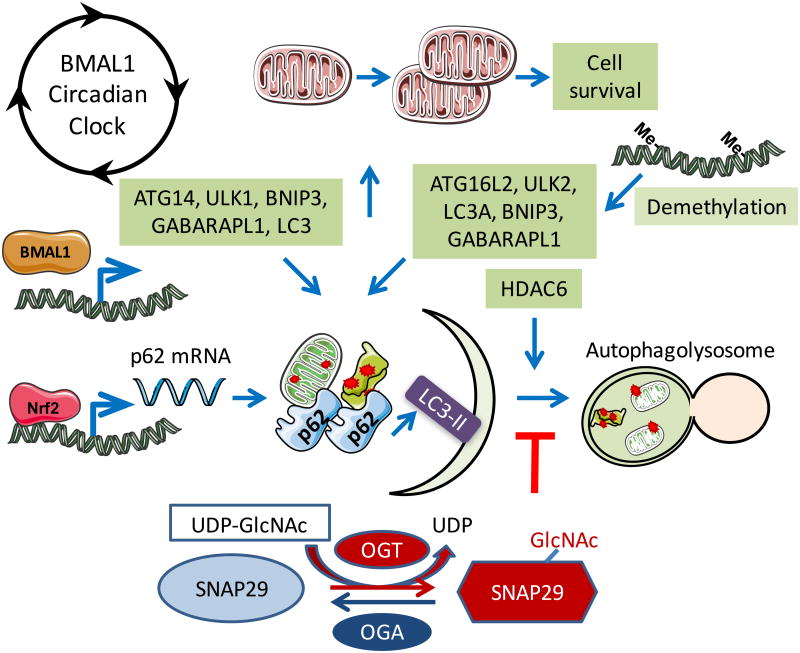
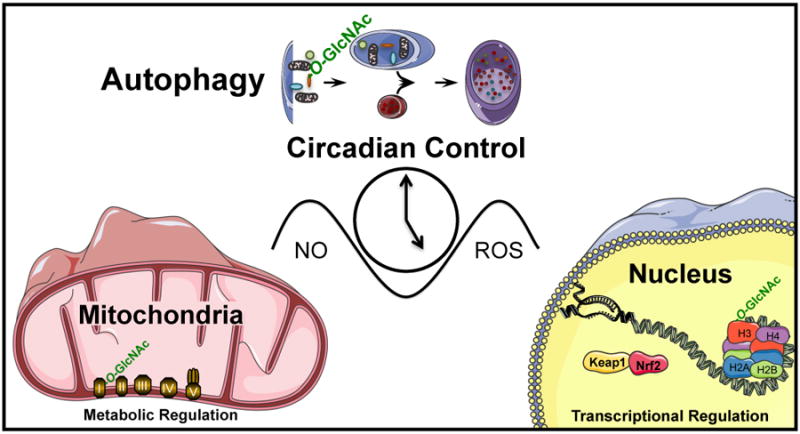
Understanding molecular mechanisms that underlie the recent emergence of metabolic diseases such as diabetes and heart failure has revealed the need for a multi-disciplinary research integrating the key metabolic pathways which change the susceptibility to environmental or pathologic stress. At the physiological level these include the circadian control of metabolism which aligns metabolism with temporal demand. The mitochondria play an important role in integrating the redox signals and metabolic flux in response to the changing activities associated with chronobiology, exercise and diet. At the molecular level this involves dynamic post-translational modifications regulating transcription, metabolism and autophagy. In this review we will discuss different examples of mechanisms which link these processes together. An important pathway capable of linking signaling to metabolism is the post-translational modification of proteins by O-linked N-acetylglucosamine (O-GlcNAc). This is a nutrient regulated protein modification that plays an important role in impaired cellular stress responses. Circadian clocks have also emerged as critical regulators of numerous cardiometabolic processes, including glucose/lipid homeostasis, hormone secretion, redox status and cardiovascular function. Central to these pathways are the response of autophagy, bioenergetics to oxidative stress, regulated by Keap1/Nrf2 and mechanisms of metabolic control. The extension of these ideas to the emerging concept of bioenergetic health will be discussed.
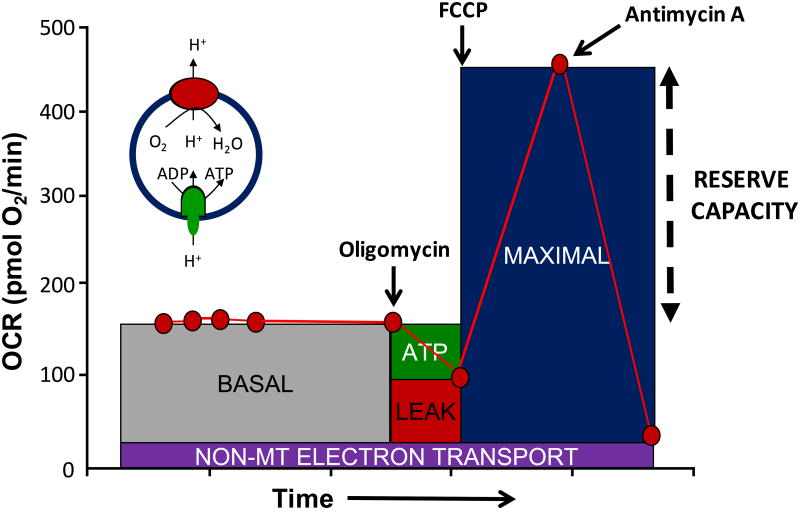
Keywords: Chronobiology; Keap1; Metabolic shift; Nrf2; Oxidative stress; Reserve capacity.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Chance B, Sies H, Boveris A. Hydroperoxide metabolism in mammalian organs. Physiol Rev. 1979;59:527–605. - PubMed
-
- Al Ghouleh I, Khoo NK, Knaus UG, Griendling KK, Touyz RM, Thannickal VJ, Barchowsky A, Nauseef WM, Kelley EE, Bauer PM, Darley-Usmar V, Shiva S, Cifuentes-Pagano E, Freeman BA, Gladwin MT, Pagano PJ. Oxidases and peroxidases in cardiovascular and lung disease: new concepts in reactive oxygen species signaling. Free radical biology & medicine. 2011;51:1271–1288. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
