

Wolbachia infections in natural Anopheles populations affect egg laying and negatively correlate with Plasmodium development
- PMID: 27243367
- PMCID: PMC4895022
- DOI: 10.1038/ncomms11772
Wolbachia infections in natural Anopheles populations affect egg laying and negatively correlate with Plasmodium development
Abstract
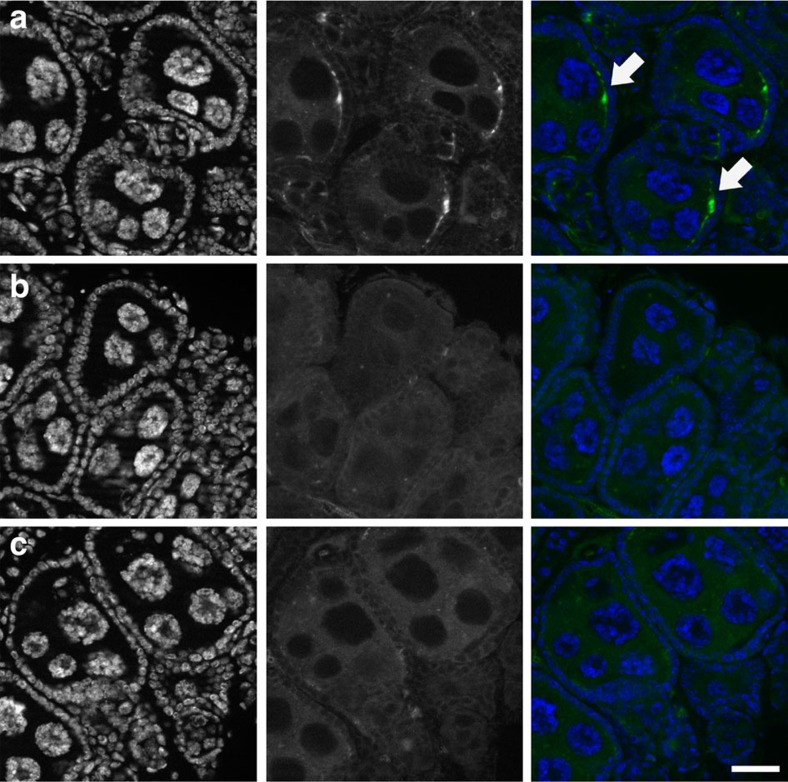
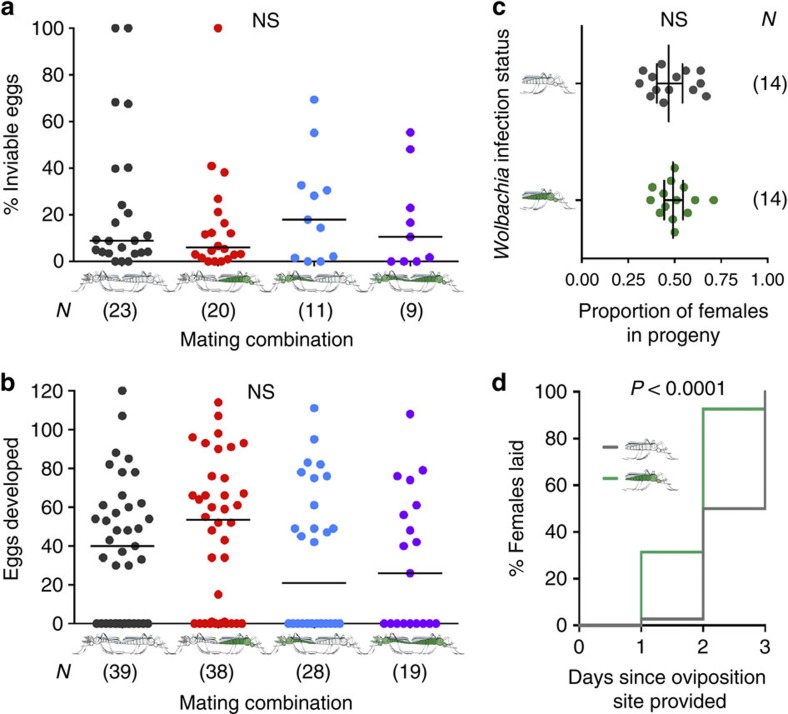
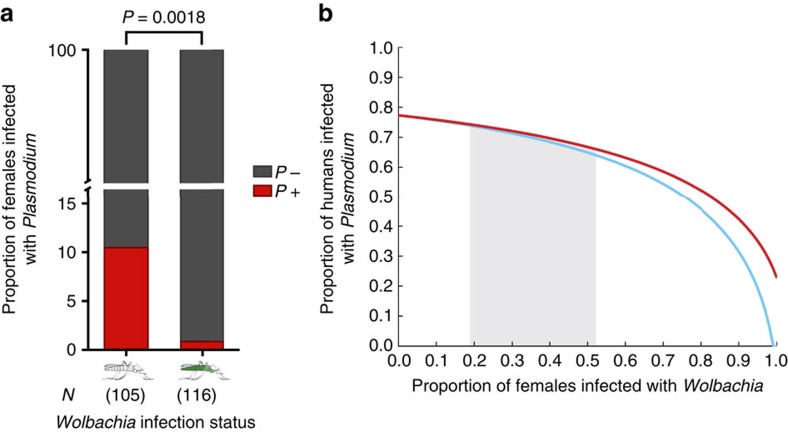
The maternally inherited alpha-proteobacterium Wolbachia has been proposed as a tool to block transmission of devastating mosquito-borne infectious diseases like dengue and malaria. Here we study the reproductive manipulations induced by a recently identified Wolbachia strain that stably infects natural mosquito populations of a major malaria vector, Anopheles coluzzii, in Burkina Faso. We determine that these infections significantly accelerate egg laying but do not induce cytoplasmic incompatibility or sex-ratio distortion, two parasitic reproductive phenotypes that facilitate the spread of other Wolbachia strains within insect hosts. Analysis of 221 blood-fed A. coluzzii females collected from houses shows a negative correlation between the presence of Plasmodium parasites and Wolbachia infection. A mathematical model incorporating these results predicts that infection with these endosymbionts may reduce malaria prevalence in human populations. These data suggest that Wolbachia may be an important player in malaria transmission dynamics in Sub-Saharan Africa.
Figures
Similar articles
-
Is Anopheles gambiae a Natural Host of Wolbachia?mBio. 2019 Jun 11;10(3):e00784-19. doi: 10.1128/mBio.00784-19. mBio. 2019. PMID: 31186318 Free PMC article.
-
The Anopheles coluzzii microbiome and its interaction with the intracellular parasite Wolbachia.Sci Rep. 2020 Aug 14;10(1):13847. doi: 10.1038/s41598-020-70745-0. Sci Rep. 2020. PMID: 32796890 Free PMC article.
-
First report of natural Wolbachia infection in the malaria mosquito Anopheles arabiensis in Tanzania.Parasit Vectors. 2018 Dec 13;11(1):635. doi: 10.1186/s13071-018-3249-y. Parasit Vectors. 2018. PMID: 30545384 Free PMC article.
-
Can Wolbachia be used to control malaria?Mem Inst Oswaldo Cruz. 2011 Aug;106 Suppl 1:212-7. doi: 10.1590/s0074-02762011000900026. Mem Inst Oswaldo Cruz. 2011. PMID: 21881776 Review.
-
The tripartite interactions between the mosquito, its microbiota and Plasmodium.Parasit Vectors. 2018 Mar 20;11(1):200. doi: 10.1186/s13071-018-2784-x. Parasit Vectors. 2018. PMID: 29558973 Free PMC article. Review.
Cited by
-
Assessing the microbiota of the snail intermediate host of trematodes, Galba truncatula.Parasit Vectors. 2024 Jan 23;17(1):31. doi: 10.1186/s13071-024-06118-7. Parasit Vectors. 2024. PMID: 38263069 Free PMC article.
-
Mosquito Trilogy: Microbiota, Immunity and Pathogens, and Their Implications for the Control of Disease Transmission.Front Microbiol. 2021 Apr 6;12:630438. doi: 10.3389/fmicb.2021.630438. eCollection 2021. Front Microbiol. 2021. PMID: 33889137 Free PMC article. Review.
-
A transgenic tool to assess Anopheles mating competitiveness in the field.Parasit Vectors. 2018 Dec 24;11(Suppl 2):651. doi: 10.1186/s13071-018-3218-5. Parasit Vectors. 2018. PMID: 30583744 Free PMC article.
-
Microbial control of arthropod-borne disease.Mem Inst Oswaldo Cruz. 2017 Feb;112(2):81-93. doi: 10.1590/0074-02760160373. Mem Inst Oswaldo Cruz. 2017. PMID: 28177042 Free PMC article. Review.
-
Holobiont perspectives on tripartite interactions among microbiota, mosquitoes, and pathogens.ISME J. 2023 Aug;17(8):1143-1152. doi: 10.1038/s41396-023-01436-7. Epub 2023 May 25. ISME J. 2023. PMID: 37231184 Free PMC article. Review.
References
-
- WHO. World Malaria Report World Health Organization (2014).
-
- Harrison G. Mosquitoes, Malaria, and Man: A History of the Hostilities Since 1880 John Murray (1978).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
