

Cellular Zinc Homeostasis Contributes to Neuronal Differentiation in Human Induced Pluripotent Stem Cells
- PMID: 27247802
- PMCID: PMC4876239
- DOI: 10.1155/2016/3760702
Cellular Zinc Homeostasis Contributes to Neuronal Differentiation in Human Induced Pluripotent Stem Cells
Abstract
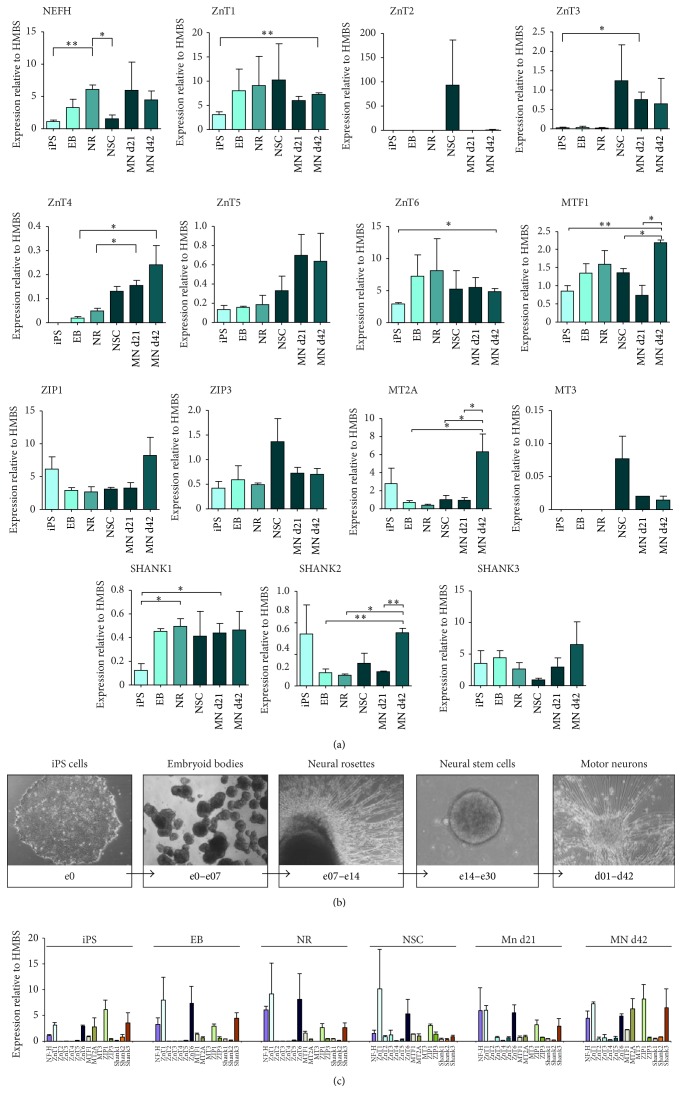
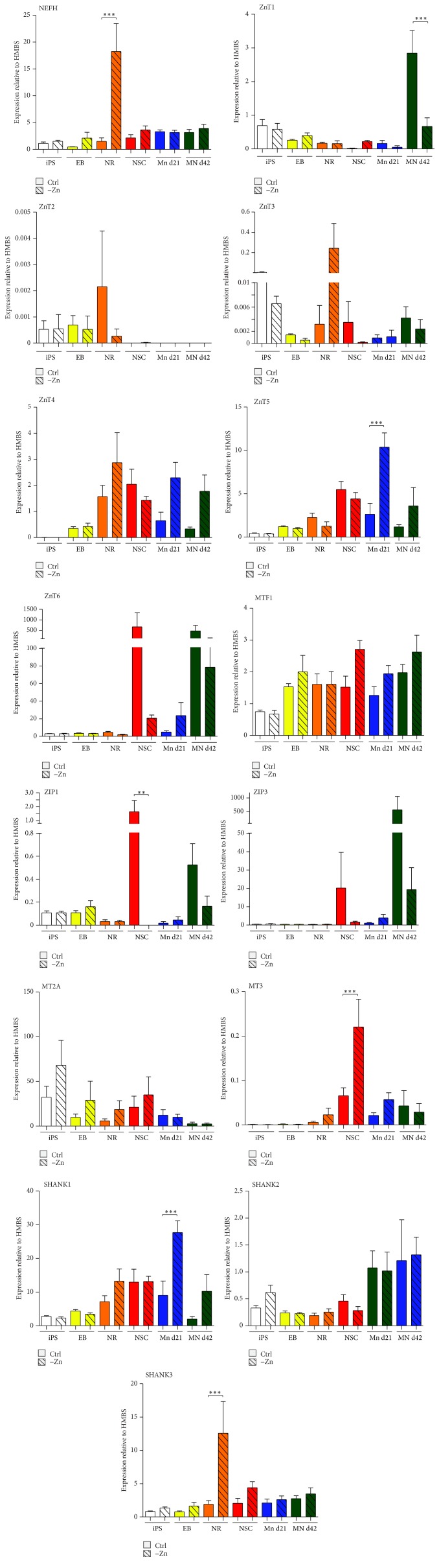
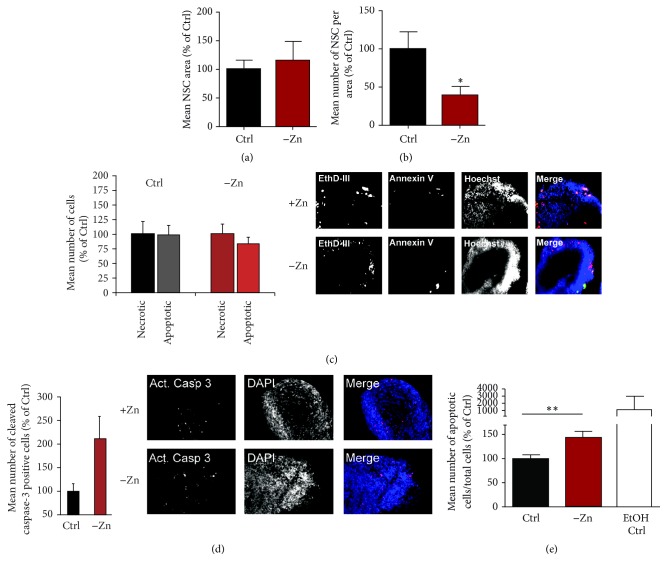
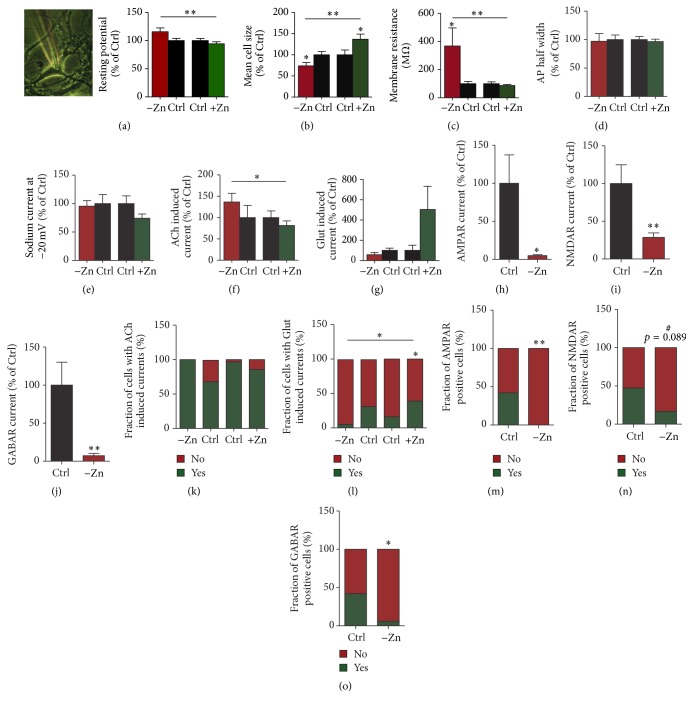
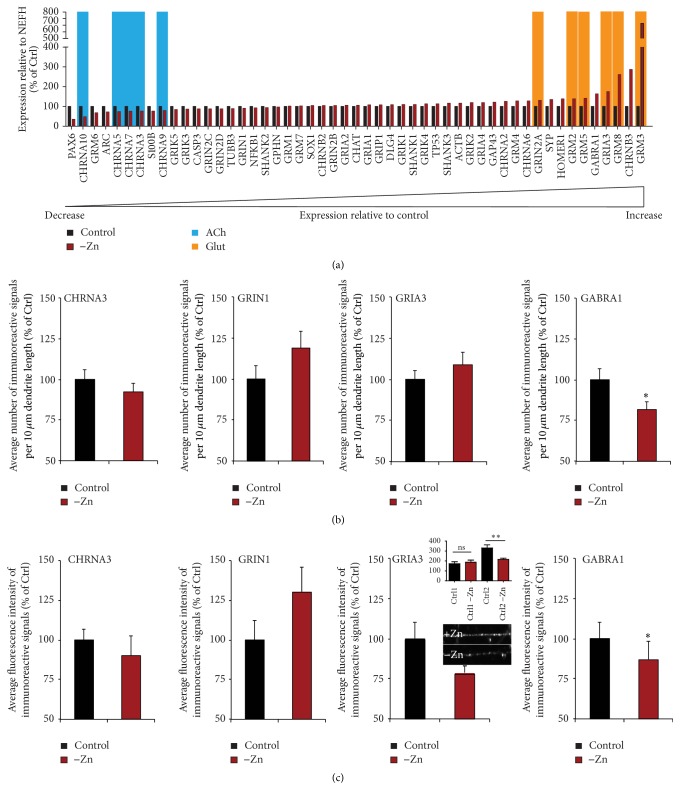
Disturbances in neuronal differentiation and function are an underlying factor of many brain disorders. Zinc homeostasis and signaling are important mediators for a normal brain development and function, given that zinc deficiency was shown to result in cognitive and emotional deficits in animal models that might be associated with neurodevelopmental disorders. One underlying mechanism of the observed detrimental effects of zinc deficiency on the brain might be impaired proliferation and differentiation of stem cells participating in neurogenesis. Thus, to examine the molecular mechanisms regulating zinc metabolism and signaling in differentiating neurons, using a protocol for motor neuron differentiation, we characterized the expression of zinc homeostasis genes during neurogenesis using human induced pluripotent stem cells (hiPSCs) and evaluated the influence of altered zinc levels on the expression of zinc homeostasis genes, cell survival, cell fate, and neuronal function. Our results show that zinc transporters are highly regulated genes during neuronal differentiation and that low zinc levels are associated with decreased cell survival, altered neuronal differentiation, and, in particular, synaptic function. We conclude that zinc deficiency in a critical time window during brain development might influence brain function by modulating neuronal differentiation.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
