

Local auxin metabolism regulates environment-induced hypocotyl elongation
- PMID: 27249562
- PMCID: PMC4849989
- DOI: 10.1038/nplants.2016.25
Local auxin metabolism regulates environment-induced hypocotyl elongation
Abstract
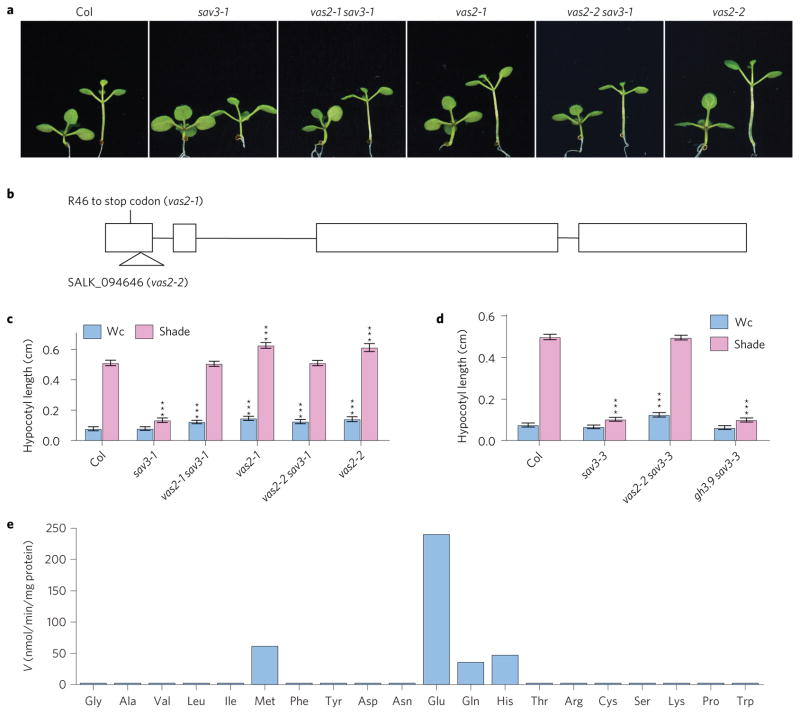
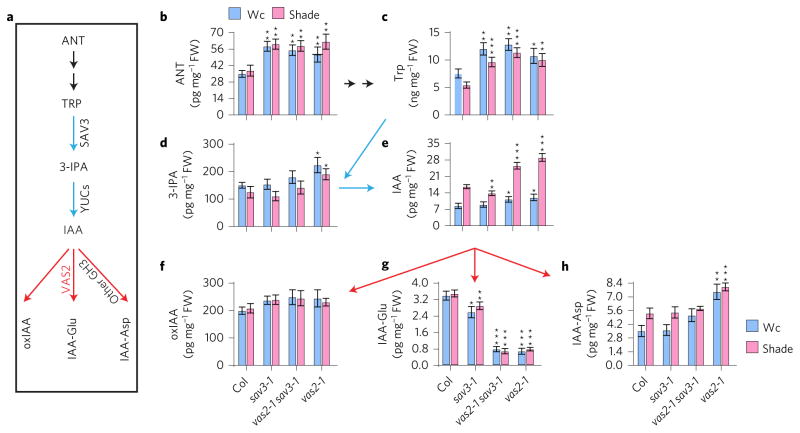
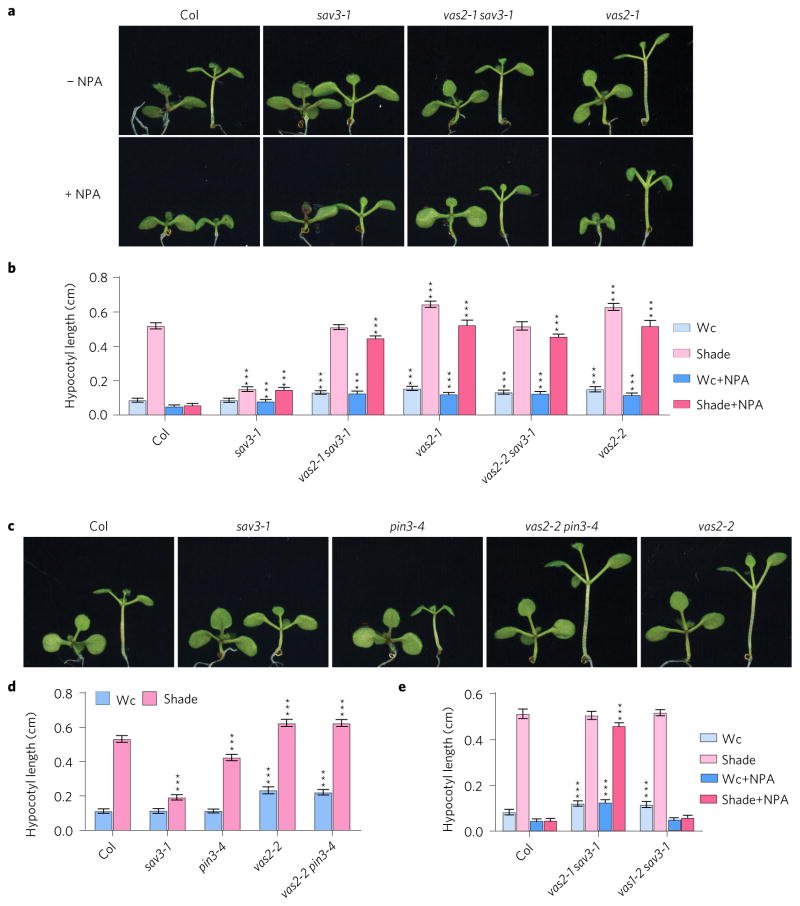
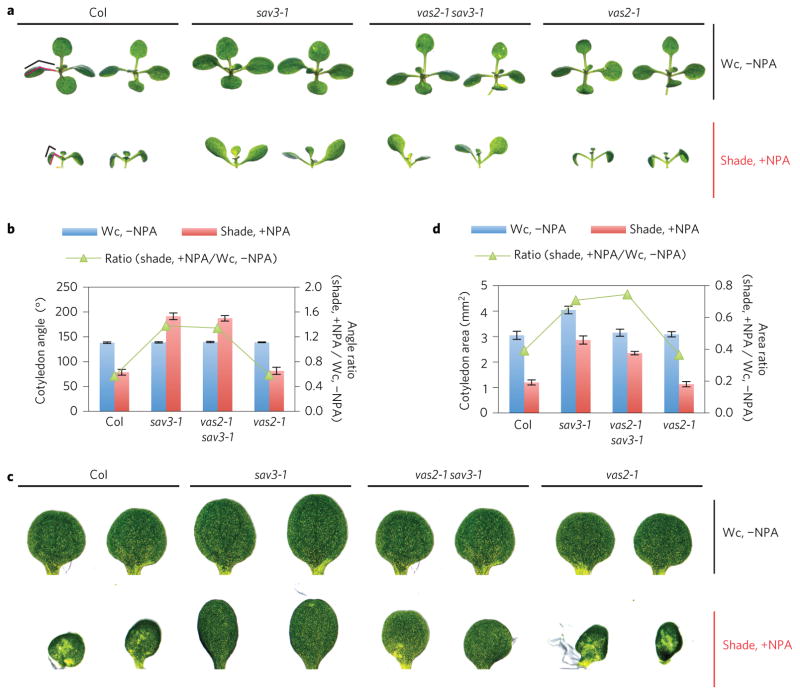
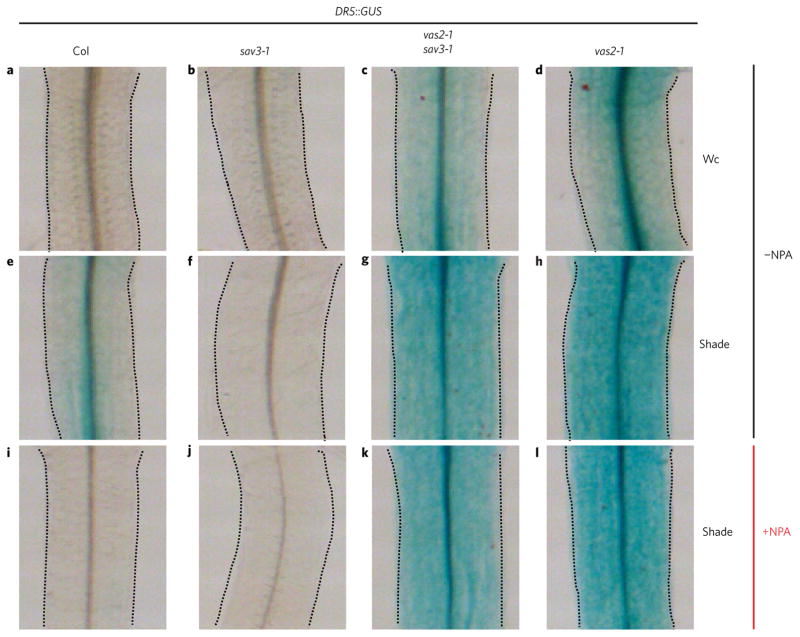
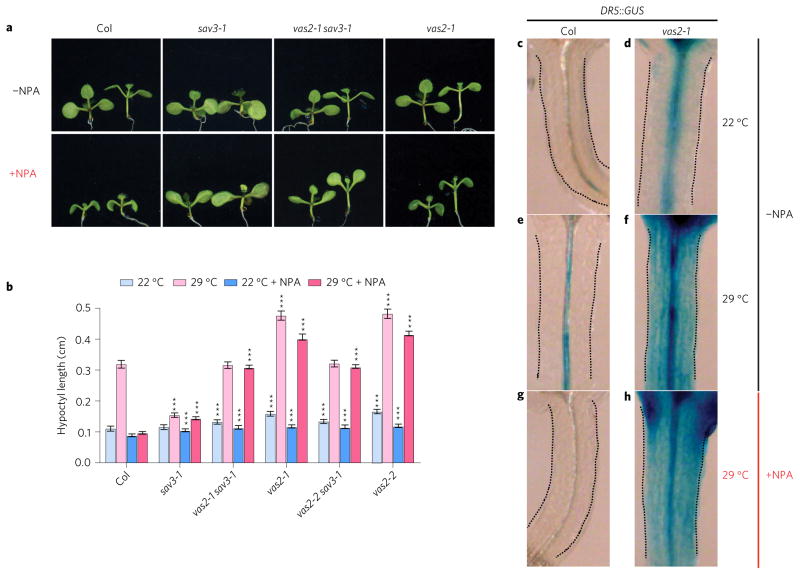
A hallmark of plants is their adaptability of size and form in response to widely fluctuating environments. The metabolism and redistribution of the phytohormone auxin play pivotal roles in establishing active auxin gradients and resulting cellular differentiation. In Arabidopsis thaliana, cotyledons and leaves synthesize indole-3-acetic acid (IAA) from tryptophan through indole-3-pyruvic acid (3-IPA) in response to vegetational shade. This newly synthesized auxin moves to the hypocotyl where it induces elongation of hypocotyl cells. Here we show that loss of function of VAS2 (IAA-amido synthetase Gretchen Hagen 3 (GH3).17) leads to increases in free IAA at the expense of IAA-Glu (IAA-glutamate) in the hypocotyl epidermis. This active IAA elicits shade- and high temperature-induced hypocotyl elongation largely independently of 3-IPA-mediated IAA biosynthesis in cotyledons. Our results reveal an unexpected capacity of local auxin metabolism to modulate the homeostasis and spatial distribution of free auxin in specialized organs such as hypocotyls in response to shade and high temperature.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
Auxin conjugation: Growing out of the shade.Nat Plants. 2016 Apr 5;2:16044. doi: 10.1038/nplants.2016.44. Nat Plants. 2016. PMID: 27249571 No abstract available.
Similar articles
-
Enhancement of hypocotyl elongation by LOV KELCH PROTEIN2 production is mediated by auxin and phytochrome-interacting factors in Arabidopsis thaliana.Plant Cell Rep. 2016 Feb;35(2):455-67. doi: 10.1007/s00299-015-1896-4. Epub 2015 Nov 25. Plant Cell Rep. 2016. PMID: 26601822
-
Inactivation of GH3.5 by COP1-mediated K63-linked ubiquitination promotes seedling hypocotyl elongation.Nat Commun. 2025 Apr 14;16(1):3541. doi: 10.1038/s41467-025-58767-6. Nat Commun. 2025. PMID: 40229271 Free PMC article.
-
Gibberellins modulate shade-induced soybean hypocotyl elongation downstream of the mutual promotion of auxin and brassinosteroids.Plant Physiol Biochem. 2020 May;150:209-221. doi: 10.1016/j.plaphy.2020.02.042. Epub 2020 Mar 3. Plant Physiol Biochem. 2020. PMID: 32155449
-
Auxin: regulation, action, and interaction.Ann Bot. 2005 Apr;95(5):707-35. doi: 10.1093/aob/mci083. Epub 2005 Mar 4. Ann Bot. 2005. PMID: 15749753 Free PMC article. Review.
-
Auxin biosynthesis and its role in plant development.Annu Rev Plant Biol. 2010;61:49-64. doi: 10.1146/annurev-arplant-042809-112308. Annu Rev Plant Biol. 2010. PMID: 20192736 Free PMC article. Review.
Cited by
-
Organ-specific transcriptional regulation by HFR1 and HY5 in response to shade in Arabidopsis.Front Plant Sci. 2024 Jul 31;15:1430639. doi: 10.3389/fpls.2024.1430639. eCollection 2024. Front Plant Sci. 2024. PMID: 39145190 Free PMC article.
-
GH3 Auxin-Amido Synthetases Alter the Ratio of Indole-3-Acetic Acid and Phenylacetic Acid in Arabidopsis.Plant Cell Physiol. 2020 Mar 1;61(3):596-605. doi: 10.1093/pcp/pcz223. Plant Cell Physiol. 2020. PMID: 31808940 Free PMC article.
-
The Roles of Auxin Biosynthesis YUCCA Gene Family in Plants.Int J Mol Sci. 2019 Dec 16;20(24):6343. doi: 10.3390/ijms20246343. Int J Mol Sci. 2019. PMID: 31888214 Free PMC article. Review.
-
Molecular breeding for improvement of photothermal adaptability in soybean.Mol Breed. 2023 Jul 24;43(8):60. doi: 10.1007/s11032-023-01406-z. eCollection 2023 Aug. Mol Breed. 2023. PMID: 37496825 Free PMC article.
-
PIF7 is a master regulator of thermomorphogenesis in shade.Nat Commun. 2022 Aug 29;13(1):4942. doi: 10.1038/s41467-022-32585-6. Nat Commun. 2022. PMID: 36038577 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
