

APOBEC4 Enhances the Replication of HIV-1
- PMID: 27249646
- PMCID: PMC4889046
- DOI: 10.1371/journal.pone.0155422
APOBEC4 Enhances the Replication of HIV-1
Abstract
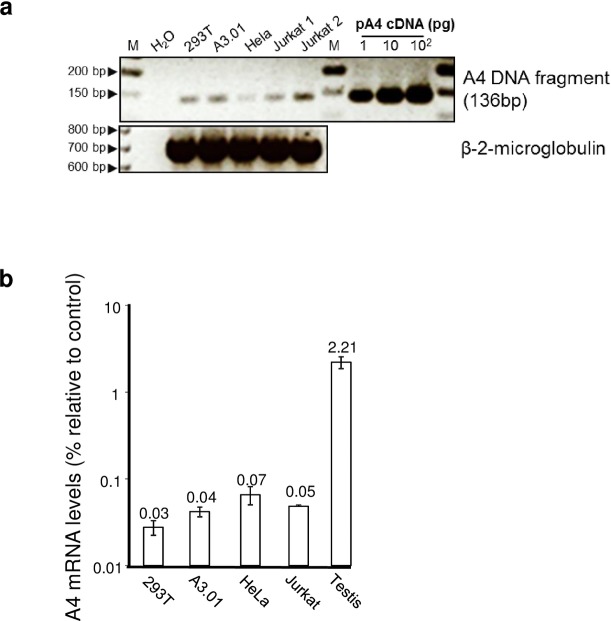
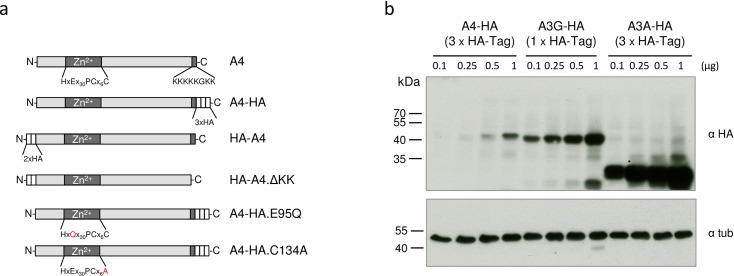
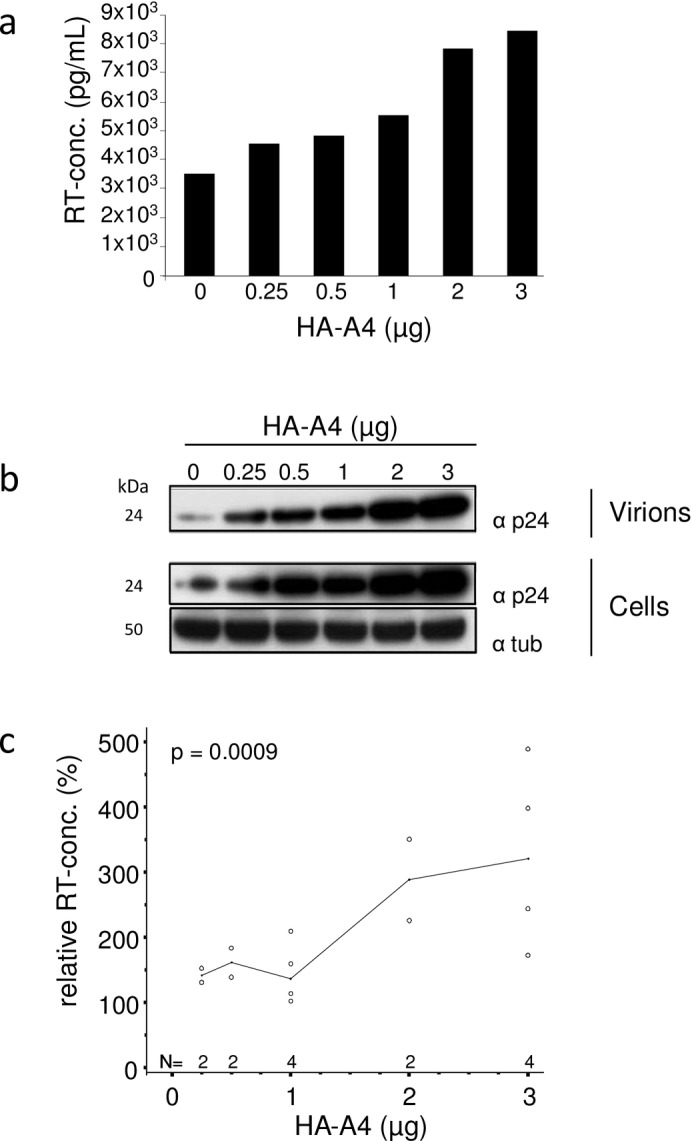
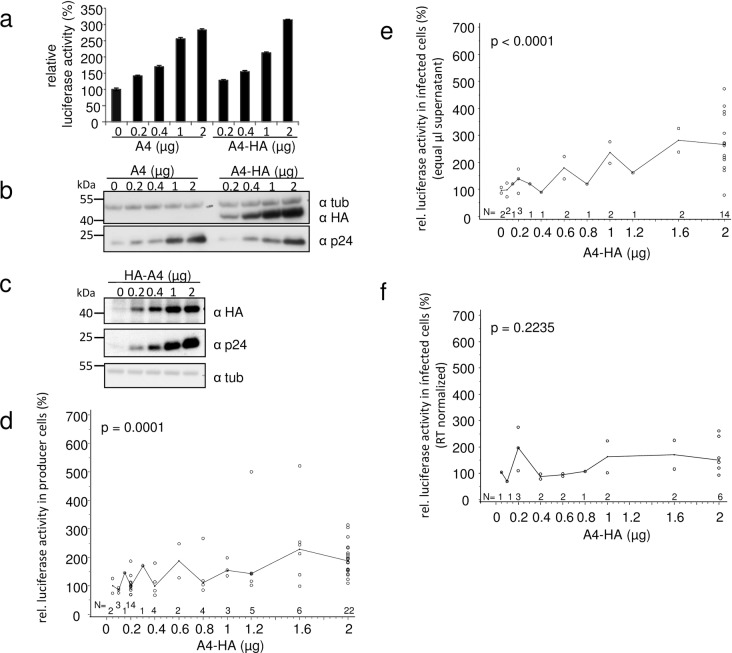
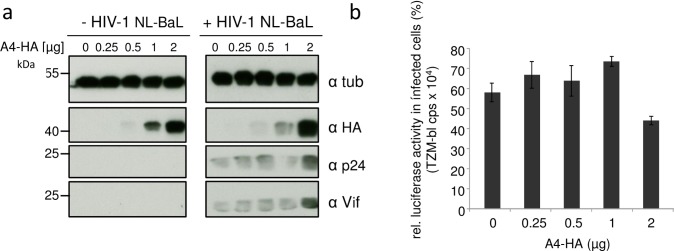
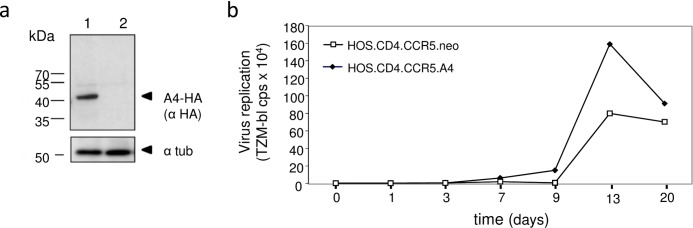
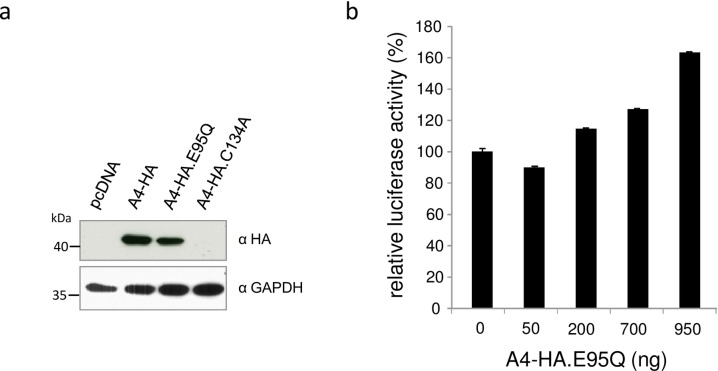
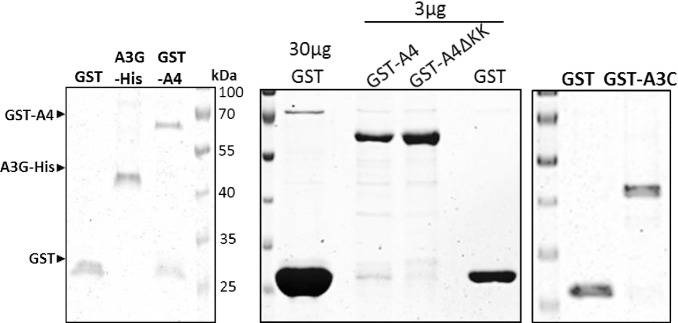
APOBEC4 (A4) is a member of the AID/APOBEC family of cytidine deaminases. In this study we found a high mRNA expression of A4 in human testis. In contrast, there were only low levels of A4 mRNA detectable in 293T, HeLa, Jurkat or A3.01 cells. Ectopic expression of A4 in HeLa cells resulted in mostly cytoplasmic localization of the protein. To test whether A4 has antiviral activity similar to that of proteins of the APOBEC3 (A3) subfamily, A4 was co-expressed in 293T cells with wild type HIV-1 and HIV-1 luciferase reporter viruses. We found that A4 did not inhibit the replication of HIV-1 but instead enhanced the production of HIV-1 in a dose-dependent manner and seemed to act on the viral LTR. A4 did not show detectable cytidine deamination activity in vitro and weakly interacted with single-stranded DNA. The presence of A4 in virus producer cells enhanced HIV-1 replication by transiently transfected A4 or stably expressed A4 in HIV-susceptible cells. APOBEC4 was capable of similarly enhancing transcription from a broad spectrum of promoters, regardless of whether they were viral or mammalian. We hypothesize that A4 may have a natural role in modulating host promoters or endogenous LTR promoters.
Conflict of interest statement
Figures

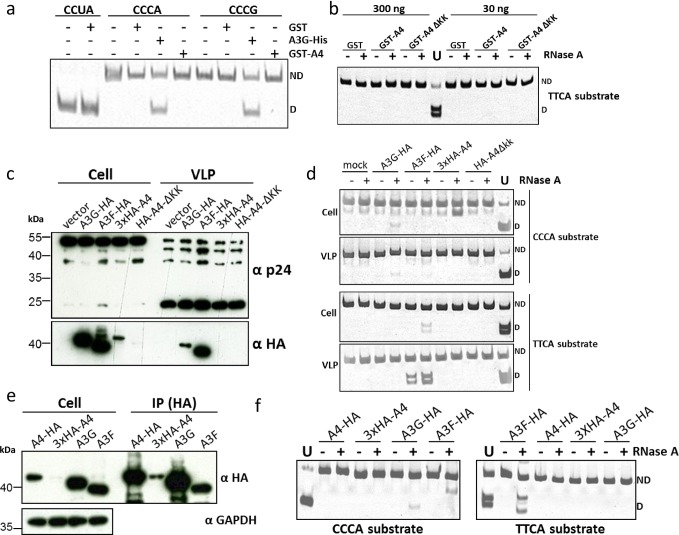
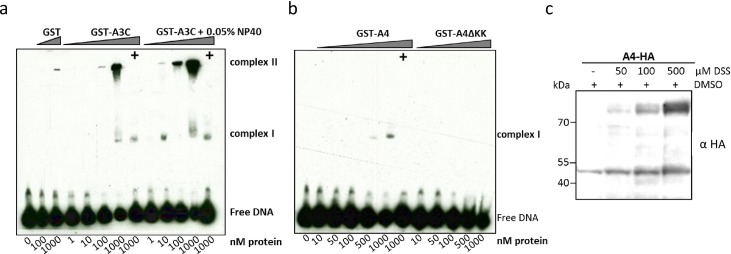
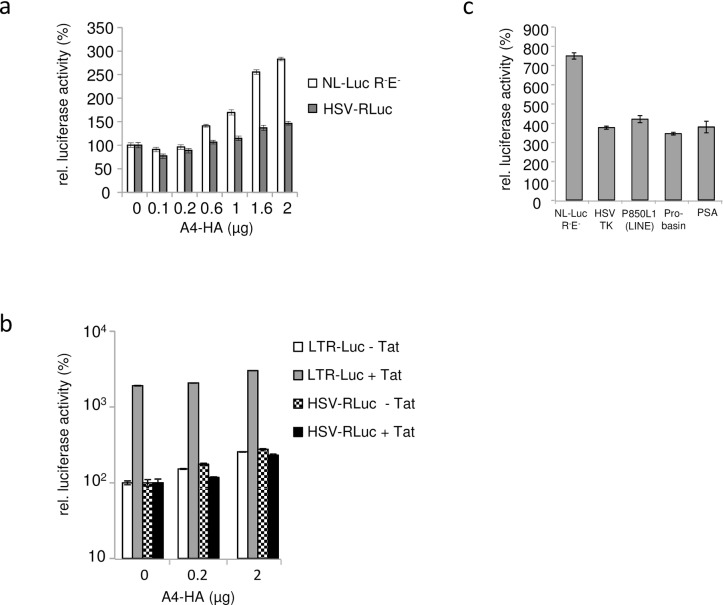
References
-
- Jarmuz A, Chester A, Bayliss J, Gisbourne J, Dunham I, Scott J, et al. An anthropoid-specific locus of orphan C to U RNA-editing enzymes on chromosome 22. Genomics. 2002;79(3):285–96. - PubMed
-
- Rogozin IB, Basu MK, Jordan IK, Pavlov YI, Koonin EV. APOBEC4, a new member of the AID/APOBEC family of polynucleotide (deoxy)cytidine deaminases predicted by computational analysis. Cell Cycle. 2005;4(9):1281–5. . - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
