

Mutagenesis of S-Adenosyl-l-Methionine-Binding Residues in Coronavirus nsp14 N7-Methyltransferase Demonstrates Differing Requirements for Genome Translation and Resistance to Innate Immunity
- PMID: 27252528
- PMCID: PMC4984653
- DOI: 10.1128/JVI.00542-16
Mutagenesis of S-Adenosyl-l-Methionine-Binding Residues in Coronavirus nsp14 N7-Methyltransferase Demonstrates Differing Requirements for Genome Translation and Resistance to Innate Immunity
Abstract
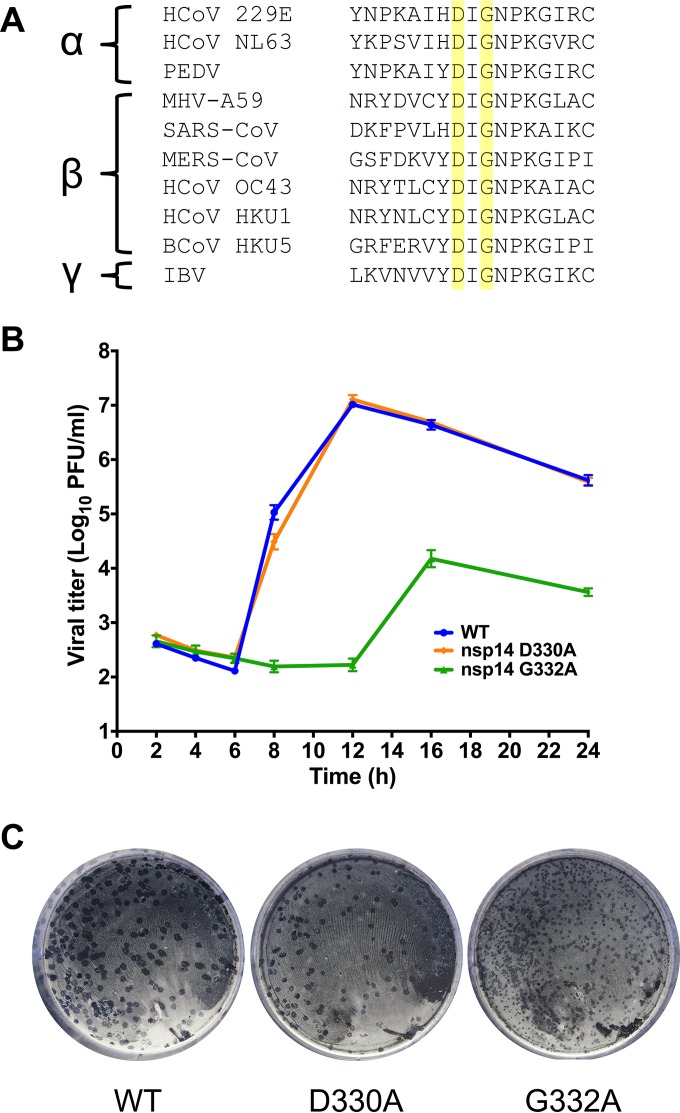
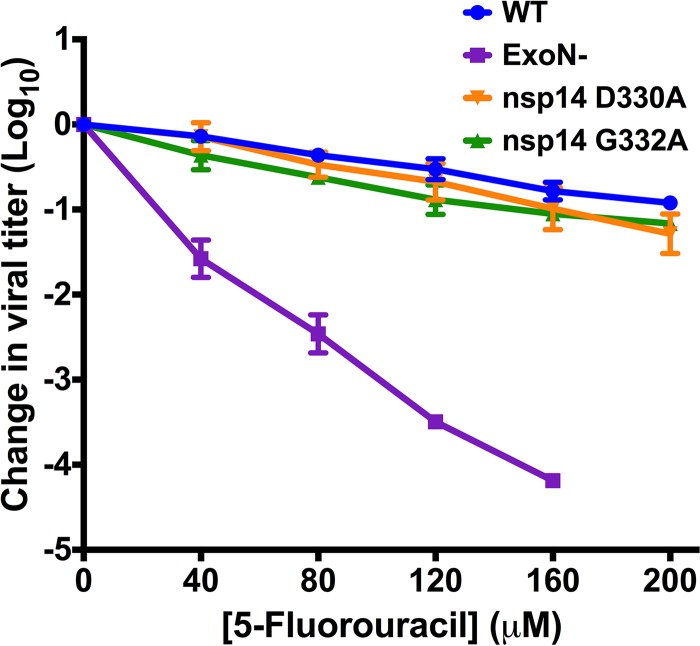
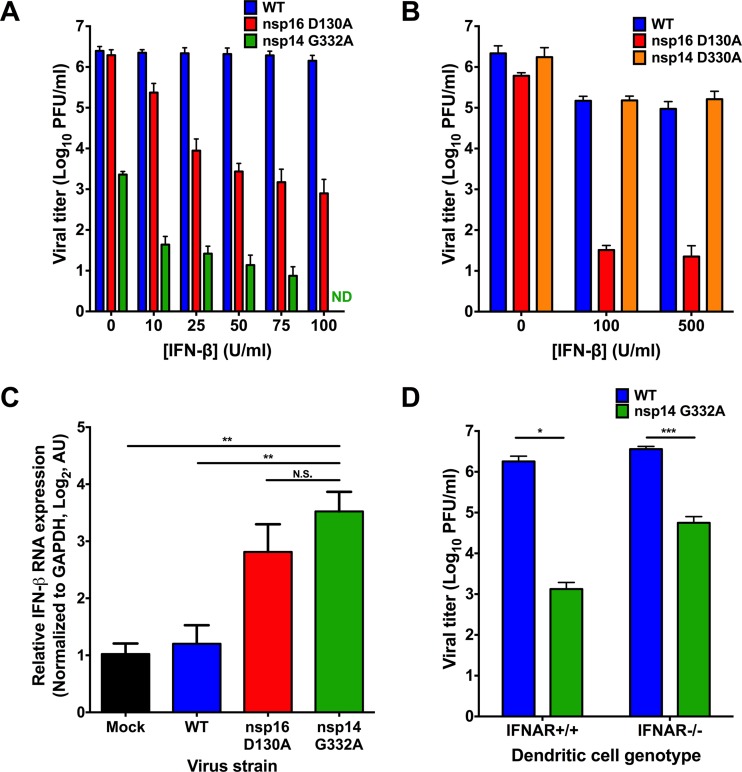
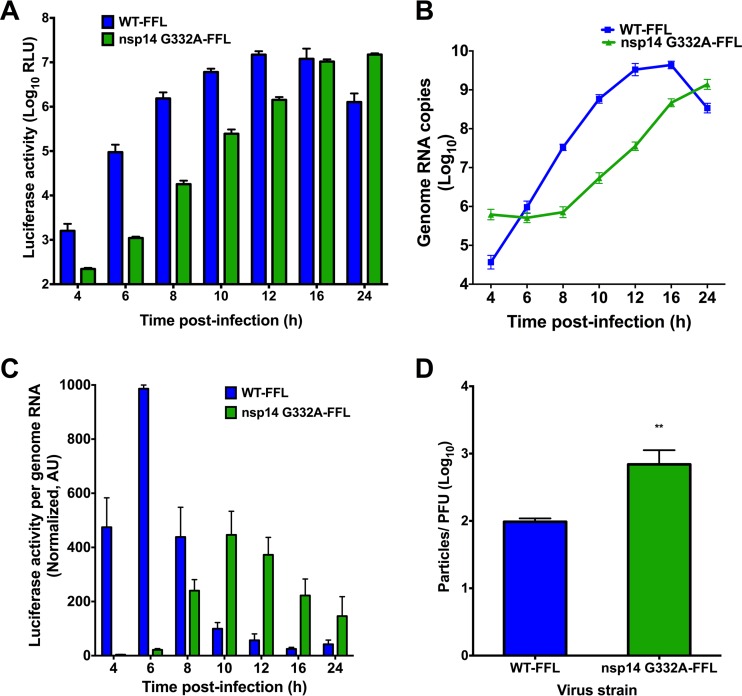
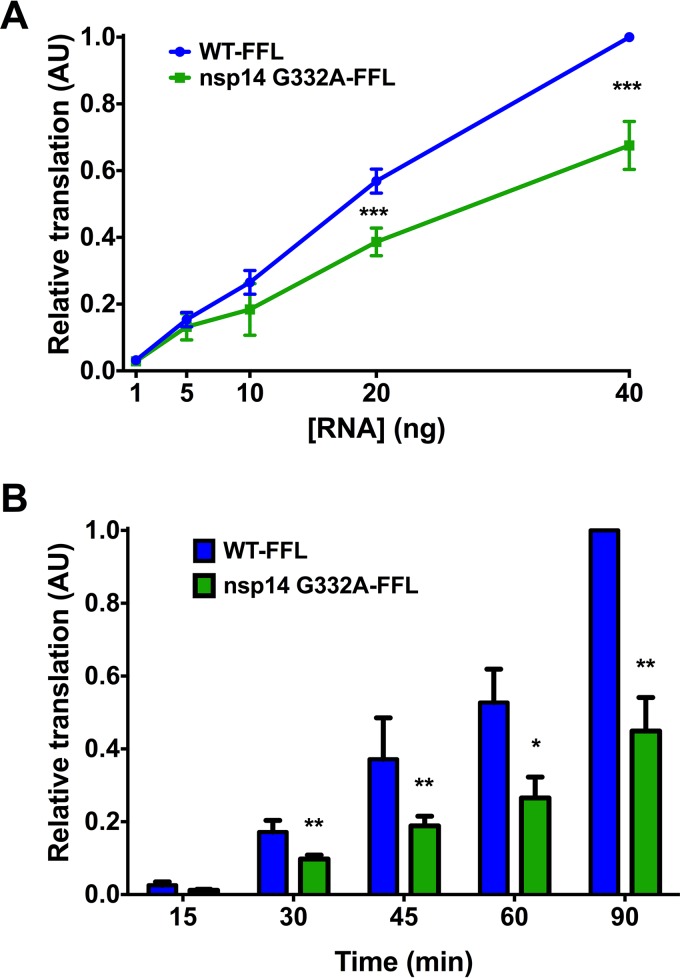
Eukaryotic mRNAs possess a methylated 5'-guanosine cap that is required for RNA stability, efficient translation, and protection from cell-intrinsic defenses. Many viruses use 5' caps or other mechanisms to mimic a cap structure to limit detection of viral RNAs by intracellular innate sensors and to direct efficient translation of viral proteins. The coronavirus (CoV) nonstructural protein 14 (nsp14) is a multifunctional protein with N7-methyltransferase (N7-MTase) activity. The highly conserved S-adenosyl-l-methionine (SAM)-binding residues of the DxG motif are required for nsp14 N7-MTase activity in vitro However, the requirement for CoV N7-MTase activity and the importance of the SAM-binding residues during viral replication have not been determined. Here, we engineered mutations in murine hepatitis virus (MHV) nsp14 N7-MTase at residues D330 and G332 and determined the effects of these mutations on viral replication, sensitivity to mutagen, inhibition by type I interferon (IFN), and translation efficiency. Virus encoding a G332A substitution in nsp14 displayed delayed replication kinetics and decreased peak titers relative to wild-type (WT) MHV. In addition, replication of nsp14 G332A virus was diminished following treatment of cells with IFN-β, and nsp14 G332A genomes were translated less efficiently both in vitro and during viral infection. In contrast, substitution of alanine at MHV nsp14 D330 did not affect viral replication, sensitivity to mutagen, or inhibition by IFN-β compared to WT MHV. Our results demonstrate that the conserved MHV N7-MTase SAM-binding-site residues are not required for MHV viability and suggest that the determinants of CoV N7-MTase activity differ in vitro and during virus infection.
Importance: Human coronaviruses, most notably severe acute respiratory syndrome (SARS)-CoV and Middle East respiratory syndrome (MERS)-CoV, cause severe and lethal human disease. Since specific antiviral therapies are not available for the treatment of human coronavirus infections, it is essential to understand the functions of conserved CoV proteins in viral replication. Here, we show that substitution of alanine at G332 in the N7-MTase domain of nsp14 impairs viral replication, enhances sensitivity to the innate immune response, and reduces viral RNA translation efficiency. Our data support the idea that coronavirus RNA capping could be targeted for development of antiviral therapeutics.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
