

Decision Making and Sequential Sampling from Memory
- PMID: 27253447
- PMCID: PMC4891701
- DOI: 10.1016/j.neuron.2016.04.036
Decision Making and Sequential Sampling from Memory
Abstract
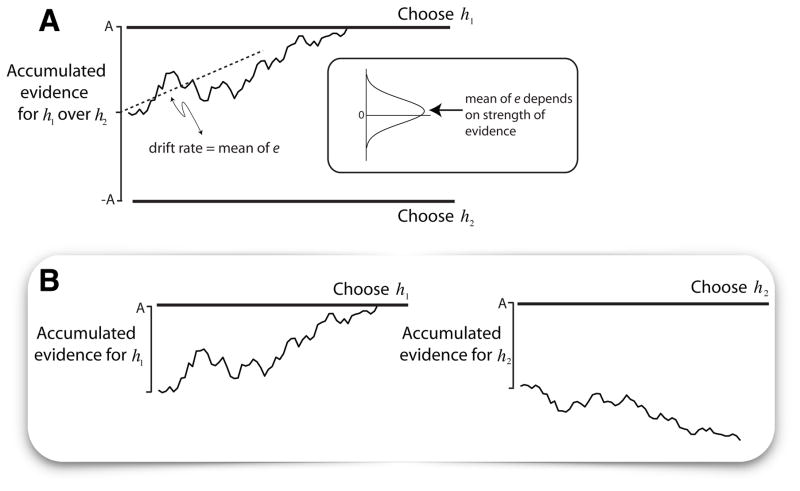
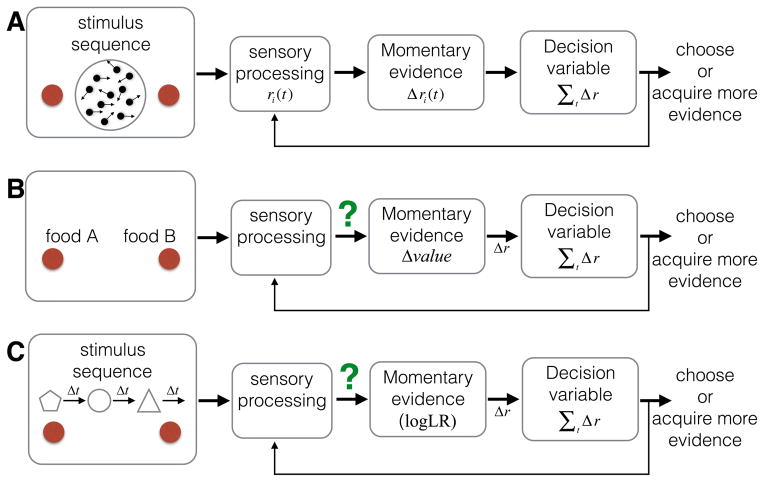
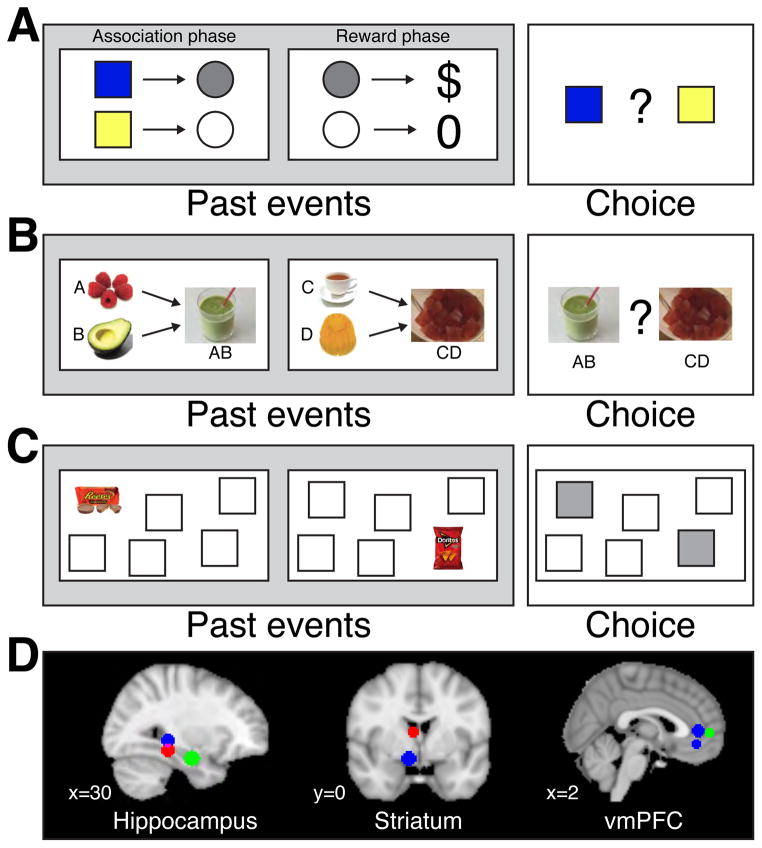
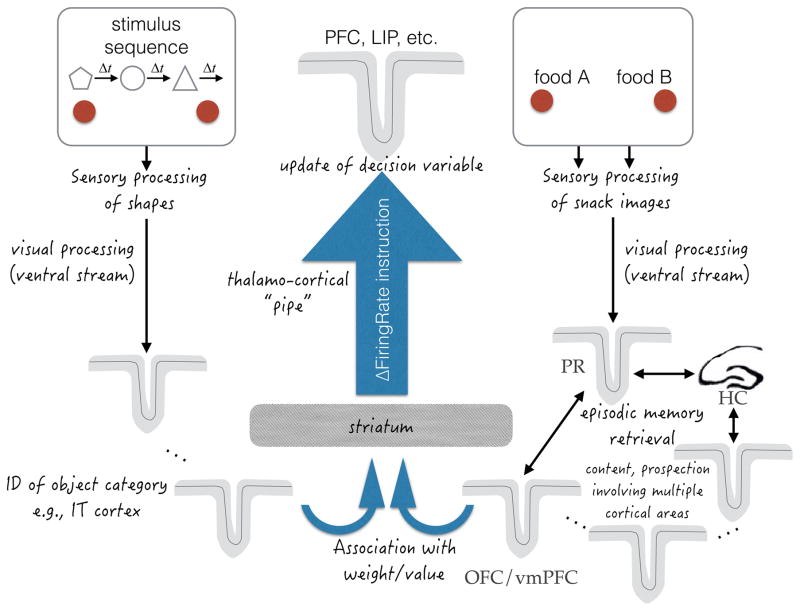
Decisions take time, and as a rule more difficult decisions take more time. But this only raises the question of what consumes the time. For decisions informed by a sequence of samples of evidence, the answer is straightforward: more samples are available with more time. Indeed, the speed and accuracy of such decisions are explained by the accumulation of evidence to a threshold or bound. However, the same framework seems to apply to decisions that are not obviously informed by sequences of evidence samples. Here, we proffer the hypothesis that the sequential character of such tasks involves retrieval of evidence from memory. We explore this hypothesis by focusing on value-based decisions and argue that mnemonic processes can account for regularities in choice and decision time. We speculate on the neural mechanisms that link sampling of evidence from memory to circuits that represent the accumulated evidence bearing on a choice. We propose that memory processes may contribute to a wider class of decisions that conform to the regularities of choice-reaction time predicted by the sequential sampling framework.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Agranov M, Ortoleva P. Stochastic Choice and Preferences for Randomization. JPE. forthcoming.
-
- Andersen RA, Cui H. Intention, action planning, and decision making in parietal-frontal circuits. Neuron. 2009;63:568–583. - PubMed
-
- Anderson JR, Bothell D, Byrne MD, Douglass S, Lebiere C, Qin Y. An Integrated Theory of the Mind. Psychological Review. 2004:1036–1060. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
