

Vitamin D Deficiency Accelerates Coronary Artery Disease Progression in Swine
- PMID: 27255724
- PMCID: PMC4965317
- DOI: 10.1161/ATVBAHA.116.307586
Vitamin D Deficiency Accelerates Coronary Artery Disease Progression in Swine
Abstract
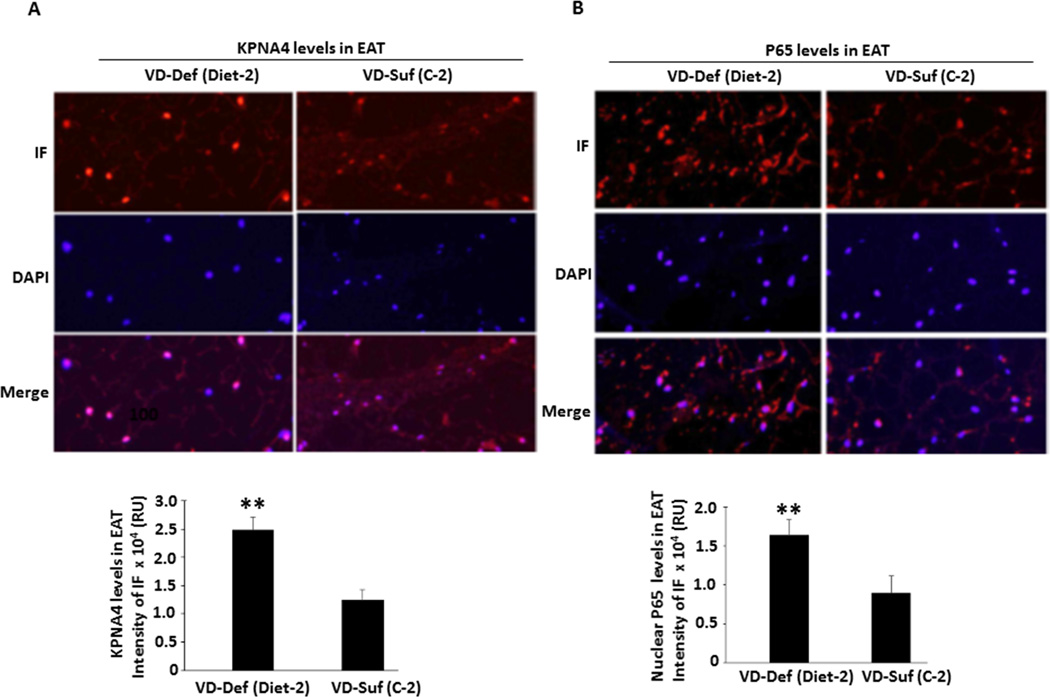
Objective: The role of vitamin D deficiency in coronary artery disease (CAD) progression is uncertain. Chronic inflammation in epicardial adipose tissue (EAT) has been implicated in the pathogenesis of CAD. However, the molecular mechanism underlying vitamin D deficiency-enhanced inflammation in the EAT of diseased coronary arteries remains unknown. We examined a mechanistic link between 1,25-dihydroxyvitamin D-mediated suppression of nuclear factor-κB (NF-κB) transporter, karyopherin α4 (KPNA4) expression and NF-κB activation in preadipocytes. Furthermore, we determined whether vitamin D deficiency accelerates CAD progression by increasing KPNA4 and nuclear NF-κB levels in EAT.
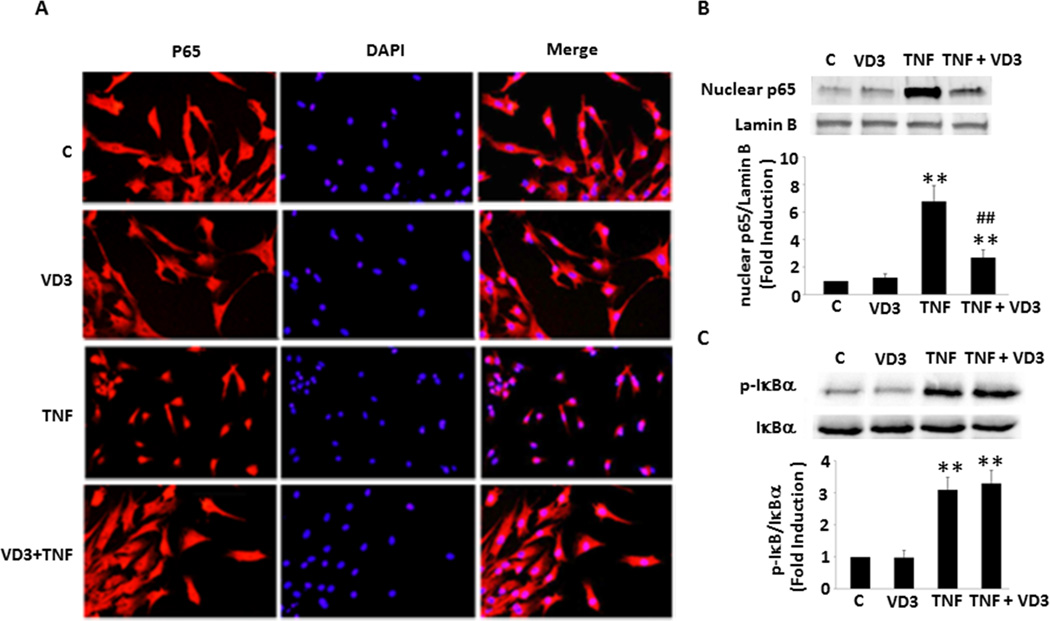
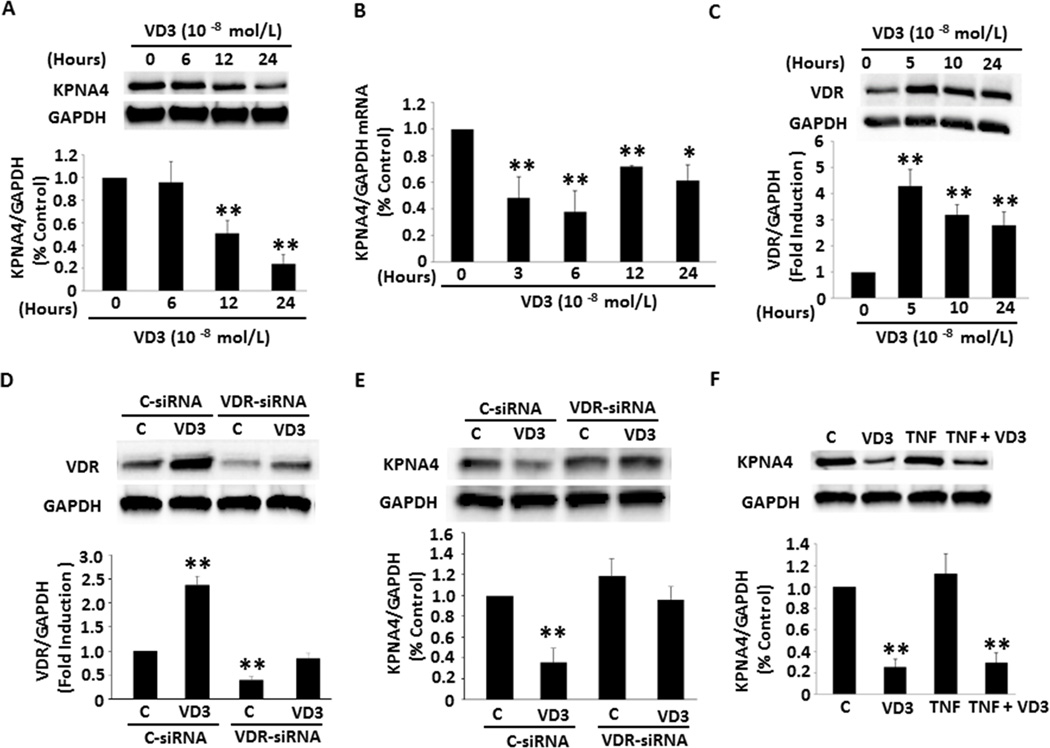
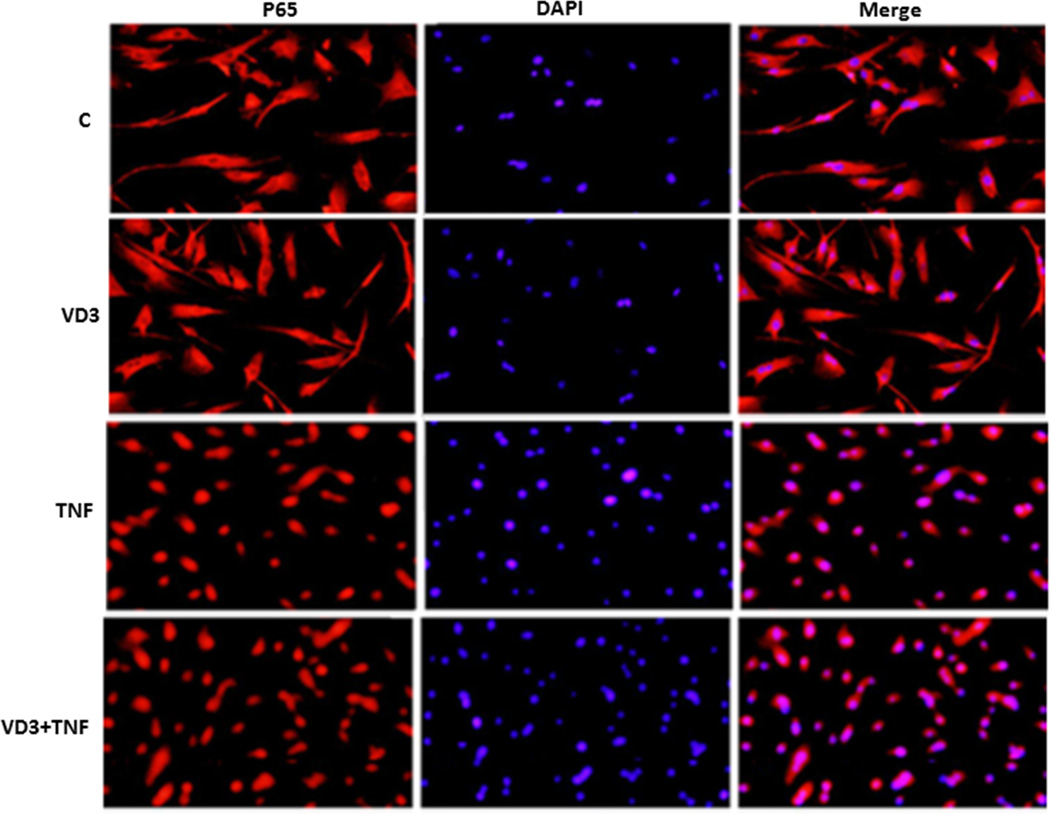
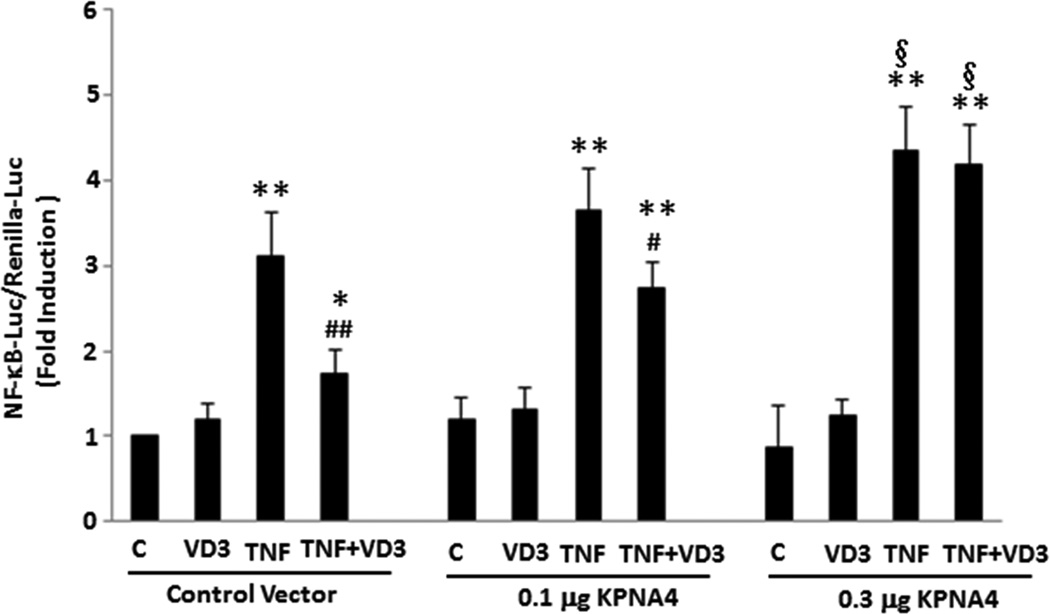
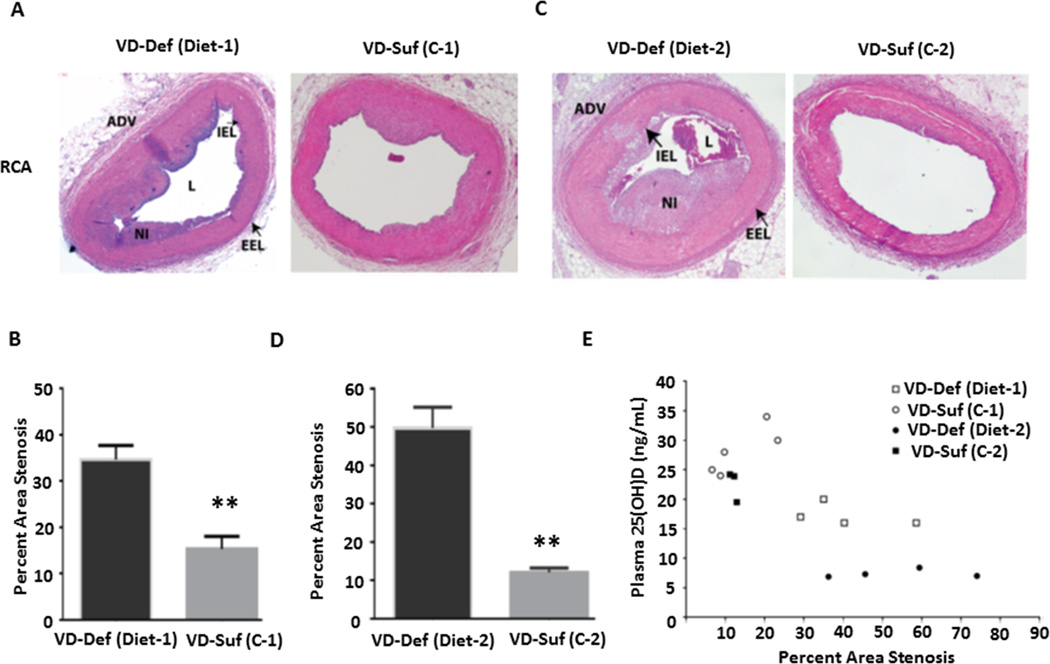
Approach and results: Nuclear protein levels were detected by immunofluorescence and Western blot. Exogenous KPNA4 was transported into cells by a transfection approach and constituted lentiviral vector. Swine were administered vitamin D-deficient or vitamin D-sufficient hypercholesterolemic diet. After 1 year, the histopathology of coronary arteries and nuclear protein expression of EAT were assessed. 1,25-dihydroxyvitamin D inhibited NF-κB activation and reduced KPNA4 levels through increased vitamin D receptor expression. Exogenous KPNA4 rescued 1,25-dihydroxyvitamin D-dependent suppression of NF-κB nuclear translocation and activation. Vitamin D deficiency caused extensive CAD progression and advanced atherosclerotic plaques, which are linked to increased KPNA4 and nuclear NF-κB levels in the EAT.
Conclusions: 1,25-dihydroxyvitamin D attenuates NF-κB activation by targeting KPNA4. Vitamin D deficiency accelerates CAD progression at least, in part, through enhanced chronic inflammation of EAT by upregulation of KPNA4, which enhances NF-κB activation. These novel findings provide mechanistic evidence that vitamin D supplementation could be beneficial for the prevention and treatment of CAD.
Keywords: 1,25-dihydroxyvitamin D; KPNA4; atherosclerosis; coronary artery disease; vitamin D.
© 2016 American Heart Association, Inc.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures
References
-
- Weber C, Noels H. Atherosclerosis: Current pathogenesis and therapeutic options. Nature medicine. 2011;17:1410–1422. - PubMed
-
- Iacobellis G, Malavazos AE, Corsi MM. Epicardial fat: From the biomolecular aspects to the clinical practice. The international journal of biochemistry & cell biology. 2011;43:1651–1654. - PubMed
-
- Mazurek T, Zhang L, Zalewski A, Mannion JD, Diehl JT, Arafat H, Sarov-Blat L, O'Brien S, Keiper EA, Johnson AG, Martin J, Goldstein BJ, Shi Y. Human epicardial adipose tissue is a source of inflammatory mediators. Circulation. 2003;108:2460–2466. - PubMed
-
- Verhagen SN, Visseren FL. Perivascular adipose tissue as a cause of atherosclerosis. Atherosclerosis. 2011;214:3–10. - PubMed
-
- Kim SH, Chung JH, Kwon BJ, Song SW, Choi WS. The associations of epicardial adipose tissue with coronary artery disease and coronary atherosclerosis. International heart journal. 2014;55:197–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
