

Functional role of mucosal-associated invariant T cells in HIV infection
- PMID: 27256572
- PMCID: PMC6608089
- DOI: 10.1189/jlb.4RU0216-084R
Functional role of mucosal-associated invariant T cells in HIV infection
Abstract
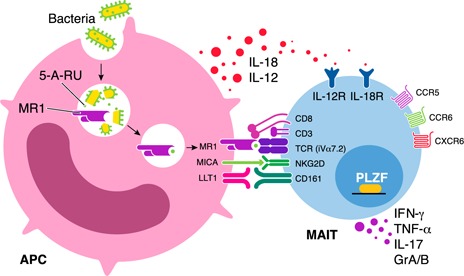
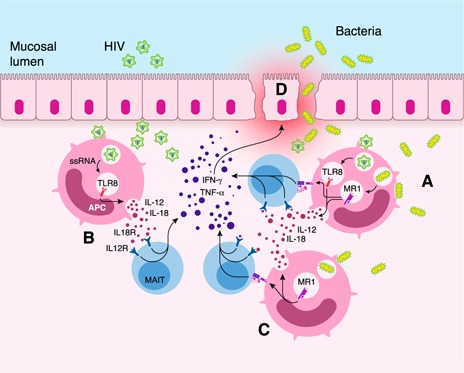
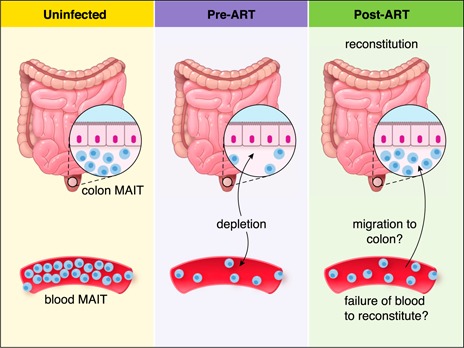
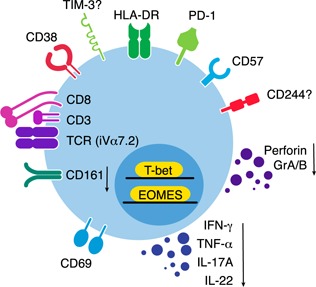
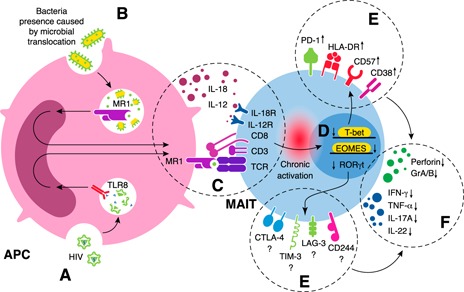
MAIT cells represent an evolutionarily conserved, MR1-restricted, innate-like cell subset that express high levels of CD161; have a canonical semi-invariant TCR iVα7.2; and may have an important role in mucosal immunity against various bacterial and fungal pathogens. Mature MAIT cells are CD161(hi)PLZF(hi)IL-18Rα(+)iVα7.2(+)γδ-CD3(+)CD8(+) T cells and occur in the peripheral blood, liver, and mucosa of humans. MAIT cells are activated by a metabolic precursor of riboflavin synthesis presented by MR1 and, therefore, respond to many bacteria and some fungi. Despite their broad antibacterial properties, their functional role in persistent viral infections is poorly understood. Although there is an increasing line of evidence portraying the depletion of MAIT cells in HIV disease, the magnitude and the potential mechanisms underlying such depletion remain unclear. Recent studies suggest that MAIT cells are vulnerable to immune exhaustion as a consequence of HIV and hepatitis C virus infections and HIV/tuberculosis coinfections. HIV infection also appears to cause functional depletion of MAIT cells resulting from abnormal expression of T-bet and EOMES, and effective ART is unable to completely salvage functional MAIT cell loss. Depletion and exhaustion of peripheral MAIT cells may affect mucosal immunity and could increase susceptibility to opportunistic infections during HIV infection. Here, we review some of the important mechanisms associated with depletion and functional loss of MAIT cells and also suggest potential immunotherapeutic strategies to restore MAIT cell functions, including the use of IL-7 to restore effector functions in HIV disease.
Keywords: CD8+ T cells; PD-1; TCR iVα7.2; cytotoxicity; exhaustion.
© Society for Leukocyte Biology.
Figures
Similar articles
-
MR1-Restricted MAIT Cells From The Human Lung Mucosal Surface Have Distinct Phenotypic, Functional, and Transcriptomic Features That Are Preserved in HIV Infection.Front Immunol. 2021 Apr 9;12:631410. doi: 10.3389/fimmu.2021.631410. eCollection 2021. Front Immunol. 2021. PMID: 33897687 Free PMC article.
-
Hyper-Expression of PD-1 Is Associated with the Levels of Exhausted and Dysfunctional Phenotypes of Circulating CD161++TCR iVα7.2+ Mucosal-Associated Invariant T Cells in Chronic Hepatitis B Virus Infection.Front Immunol. 2018 Mar 19;9:472. doi: 10.3389/fimmu.2018.00472. eCollection 2018. Front Immunol. 2018. PMID: 29616020 Free PMC article. Clinical Trial.
-
Multiple layers of heterogeneity and subset diversity in human MAIT cell responses to distinct microorganisms and to innate cytokines.Proc Natl Acad Sci U S A. 2017 Jul 3;114(27):E5434-E5443. doi: 10.1073/pnas.1705759114. Epub 2017 Jun 19. Proc Natl Acad Sci U S A. 2017. PMID: 28630305 Free PMC article.
-
MAIT, MR1, microbes and riboflavin: a paradigm for the co-evolution of invariant TCRs and restricting MHCI-like molecules?Immunogenetics. 2016 Aug;68(8):537-48. doi: 10.1007/s00251-016-0927-9. Epub 2016 Jul 8. Immunogenetics. 2016. PMID: 27393664 Review.
-
MHC class I-related molecule, MR1, and mucosal-associated invariant T cells.Immunol Rev. 2016 Jul;272(1):120-38. doi: 10.1111/imr.12423. Immunol Rev. 2016. PMID: 27319347 Review.
Cited by
-
Understanding the Role of Mucosal-Associated Invariant T-Cells in Non-human Primate Models of HIV Infection.Front Immunol. 2020 Aug 18;11:2038. doi: 10.3389/fimmu.2020.02038. eCollection 2020. Front Immunol. 2020. PMID: 33013862 Free PMC article. Review.
-
Mucosal-Associated Invariant T Cells in Regenerative Medicine.Front Immunol. 2017 Dec 1;8:1711. doi: 10.3389/fimmu.2017.01711. eCollection 2017. Front Immunol. 2017. PMID: 29250077 Free PMC article. Review.
-
Close Encounters of Lymphoid Cells and Bacteria.Front Immunol. 2016 Oct 7;7:405. doi: 10.3389/fimmu.2016.00405. eCollection 2016. Front Immunol. 2016. PMID: 27774092 Free PMC article. Review.
-
Human MAIT cells exit peripheral tissues and recirculate via lymph in steady state conditions.JCI Insight. 2018 Apr 5;3(7):e98487. doi: 10.1172/jci.insight.98487. eCollection 2018 Apr 5. JCI Insight. 2018. PMID: 29618662 Free PMC article.
-
The gut microbiome as a biomarker of differential susceptibility to SARS-CoV-2.Trends Mol Med. 2021 Dec;27(12):1115-1134. doi: 10.1016/j.molmed.2021.09.009. Epub 2021 Oct 6. Trends Mol Med. 2021. PMID: 34756546 Free PMC article. Review.
References
-
- Dusseaux, M. , Martin, E. , Serriari, N. , Péguillet, I. , Premel, V. , Louis, D. , Milder, M. , Le Bourhis, L. , Soudais, C. , Treiner, E. , Lantz, O. (2011) Human MAIT cells are xenobiotic‐resistant, tissue‐targeted, CD161hi IL‐17‐secreting T cells. Blood 117, 1250–1259. - PubMed
-
- Le Bourhis, L. , Mburu, Y. K. , Lantz, O. (2013) MAIT cells, surveyors of a new class of antigen: development and functions. Curr. Opin. Immunol. 25, 174–180. - PubMed
-
- Treiner, E. , Duban, L. , Bahram, S. , Radosavljevic, M. , Wanner, V. , Tilloy, F. , Affaticati, P. , Gilfillan, S. , Lantz, O. (2003) Selection of evolutionarily conserved mucosal‐associated invariant T cells by MR1. Nature 422, 164–169. - PubMed
-
- Reantragoon, R. , Kjer‐Nielsen, L. , Patel, O. , Chen, Z. , Illing, P. T. , Bhati, M. , Kostenko, L. , Bharadwaj, M. , Meehan, B. , Hansen, T. H. , Godfrey, D. I. , Rossjohn, J. , McCluskey, J. (2012) Structural insight into MR1‐mediated recognition of the mucosal associated invariant T cell receptor. J. Exp. Med. 209, 761–774. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
