

Distinct mechanisms regulate Cdx2 expression in the blastocyst and in trophoblast stem cells
- PMID: 27256674
- PMCID: PMC4891713
- DOI: 10.1038/srep27139
Distinct mechanisms regulate Cdx2 expression in the blastocyst and in trophoblast stem cells
Abstract
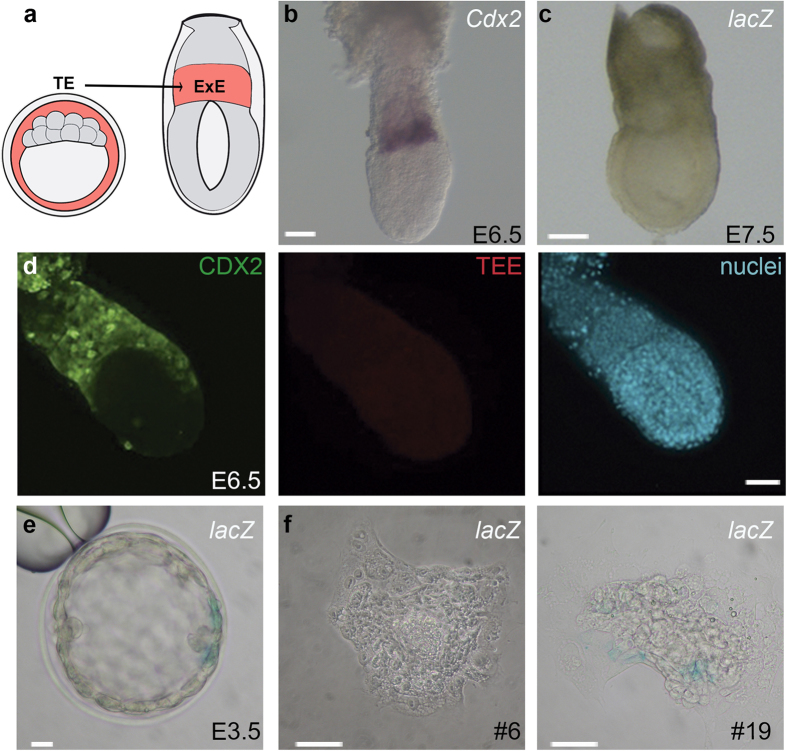
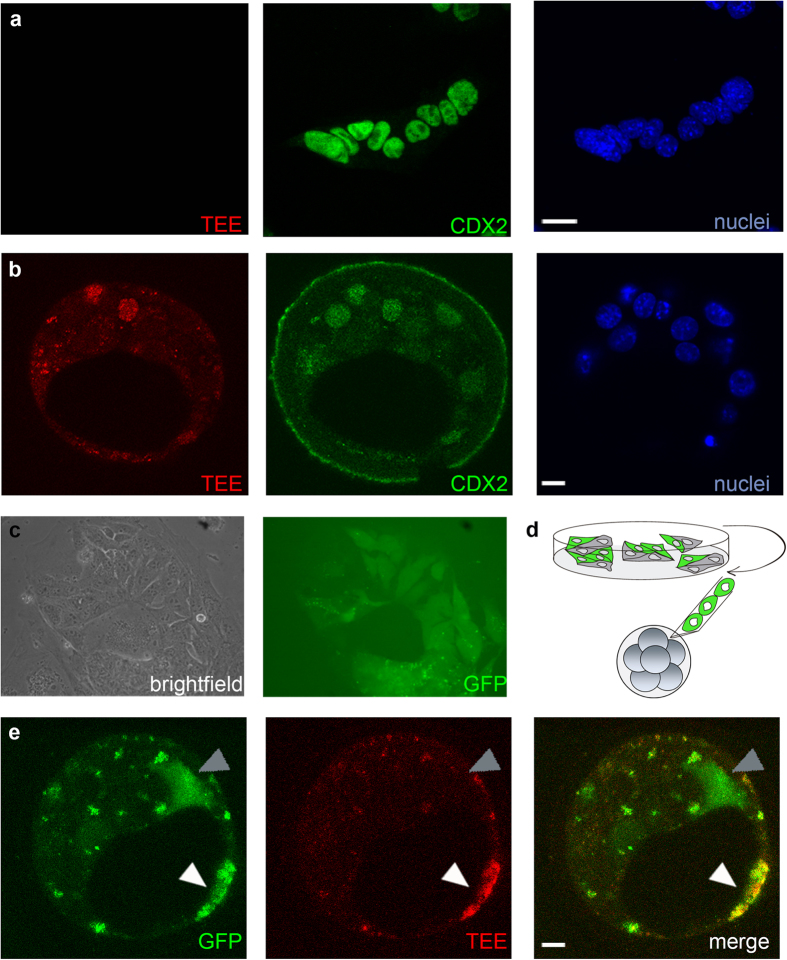
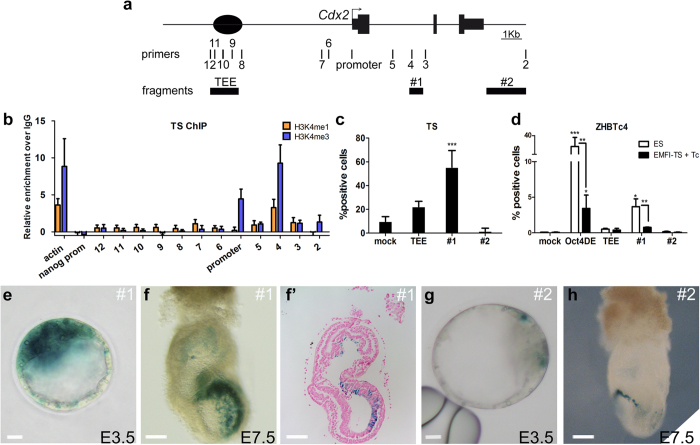
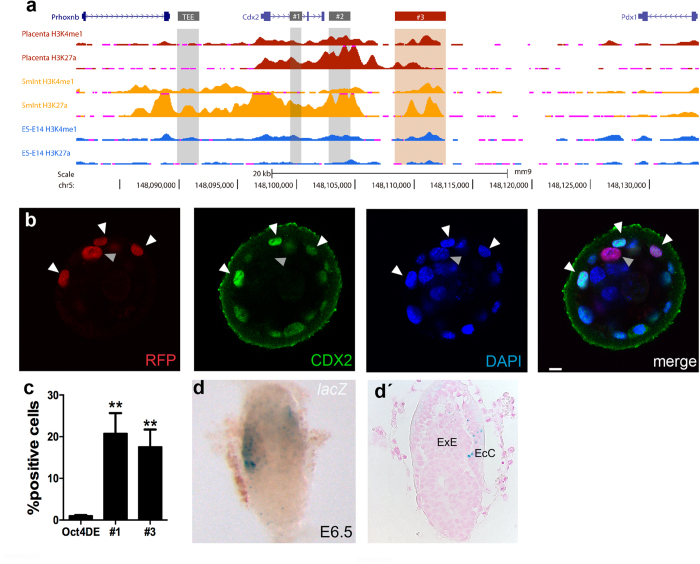
The first intercellular differences during mammalian embryogenesis arise in the blastocyst, producing the inner cell mass and the trophectoderm. The trophectoderm is the first extraembryonic tissue and does not contribute to the embryo proper, its differentiation instead forming tissues that sustain embryonic development. Crucial roles in extraembryonic differentiation have been identified for certain transcription factors, but a comprehensive picture of the regulation of this early specification is still lacking. Here, we investigated whether the regulatory mechanisms involved in Cdx2 expression in the blastocyst are also utilized in the postimplantation embryo. We analyzed an enhancer that is regulated through Hippo and Notch in the blastocyst trophectoderm, unexpectedly finding that it is inactive in the extraembryonic structures at postimplantation stages. Further analysis identified other Cdx2 regulatory elements including a stem-cell specific regulatory sequence and an element that drives reporter expression in the trophectoderm, a subset of cells in the extraembryonic region of the postimplantation embryo and in trophoblast stem cells. The cross-comparison in this study of cis-regulatory elements employed in the blastocyst, stem cell populations and the postimplantation embryo provides new insights into early mammalian development and suggests a two-step mechanism in Cdx2 regulation.
Figures
References
-
- Roper S. & Hemberger M. Defining pathways that enforce cell lineage specification in early development and stem cells. Cell Cycle 8, 1515–1525 (2009). - PubMed
-
- Yamanaka Y., Ralston A., Stephenson R. O. & Rossant J. Cell and molecular regulation of the mouse blastocyst. Dev Dyn 235, 2301–2314 (2006). - PubMed
-
- Tanaka S., Kunath T., Hadjantonakis A. K., Nagy A. & Rossant J. Promotion of trophoblast stem cell proliferation by FGF4. Science 282, 2072–2075 (1998). - PubMed
-
- Nishioka N. et al. The Hippo signaling pathway components Lats and Yap pattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass. Dev Cell 16, 398–410 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
