

Triggering the adaptive immune system with commensal gut bacteria protects against insulin resistance and dysglycemia
- PMID: 27257599
- PMCID: PMC4877664
- DOI: 10.1016/j.molmet.2016.03.004
Triggering the adaptive immune system with commensal gut bacteria protects against insulin resistance and dysglycemia
Abstract
Objective: To demonstrate that glycemia and insulin resistance are controlled by a mechanism involving the adaptive immune system and gut microbiota crosstalk.
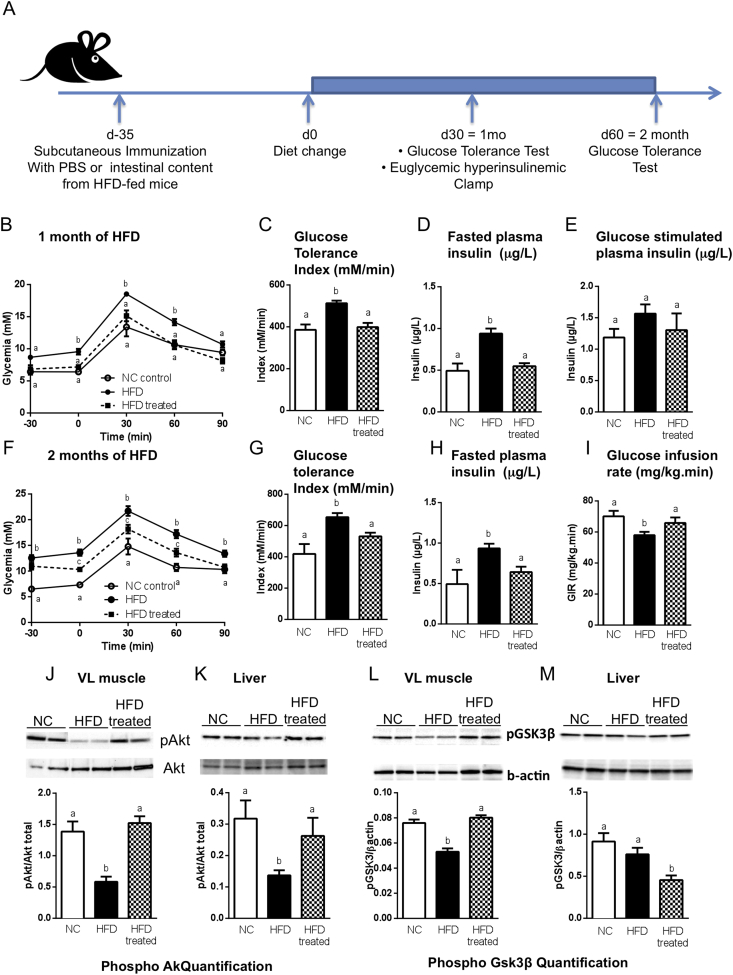
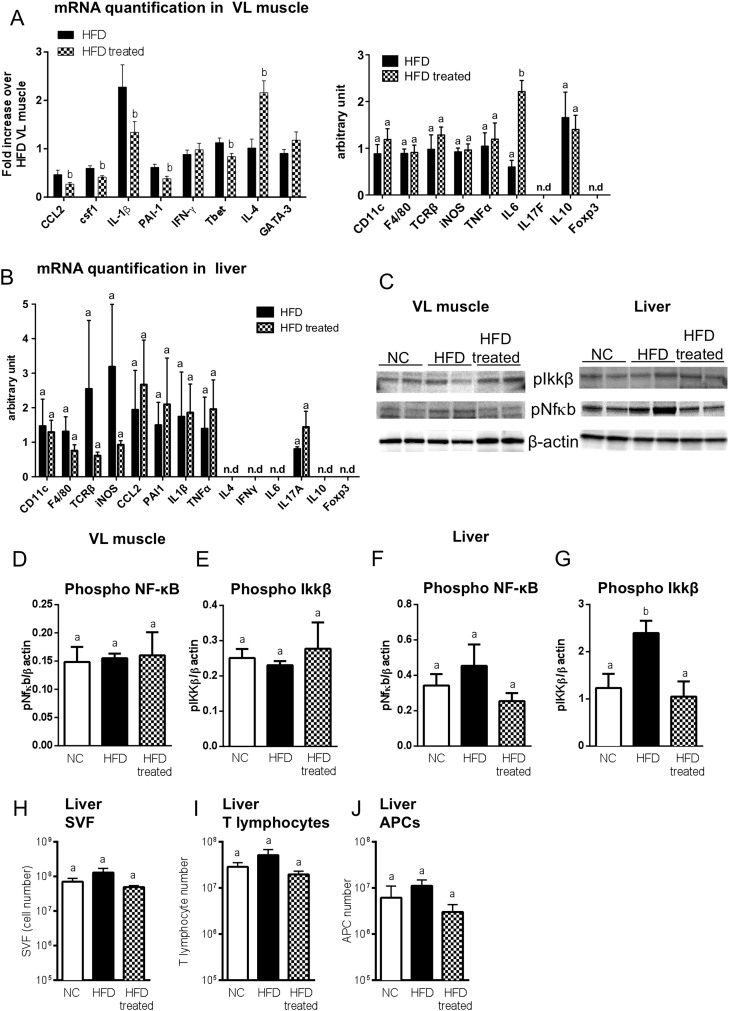
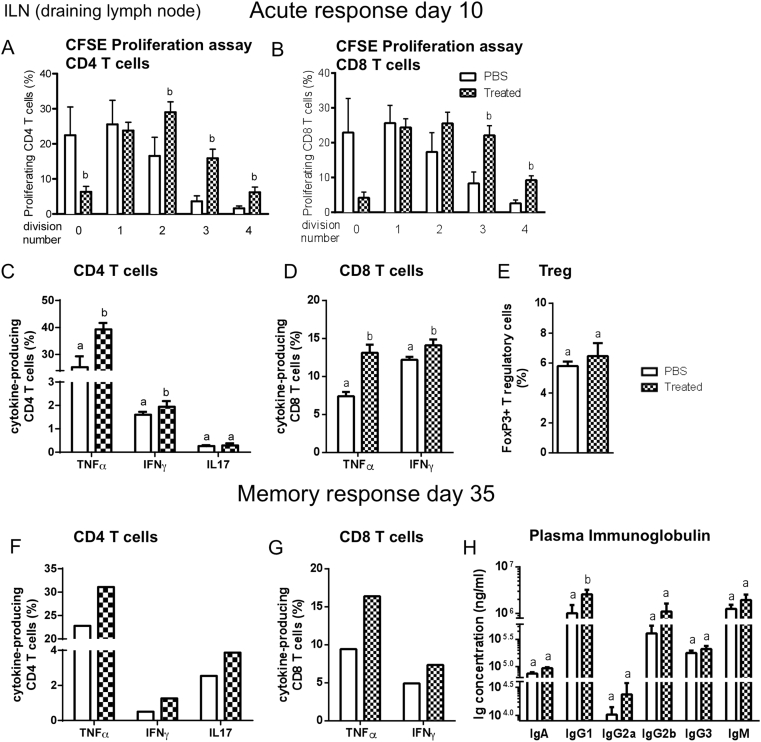
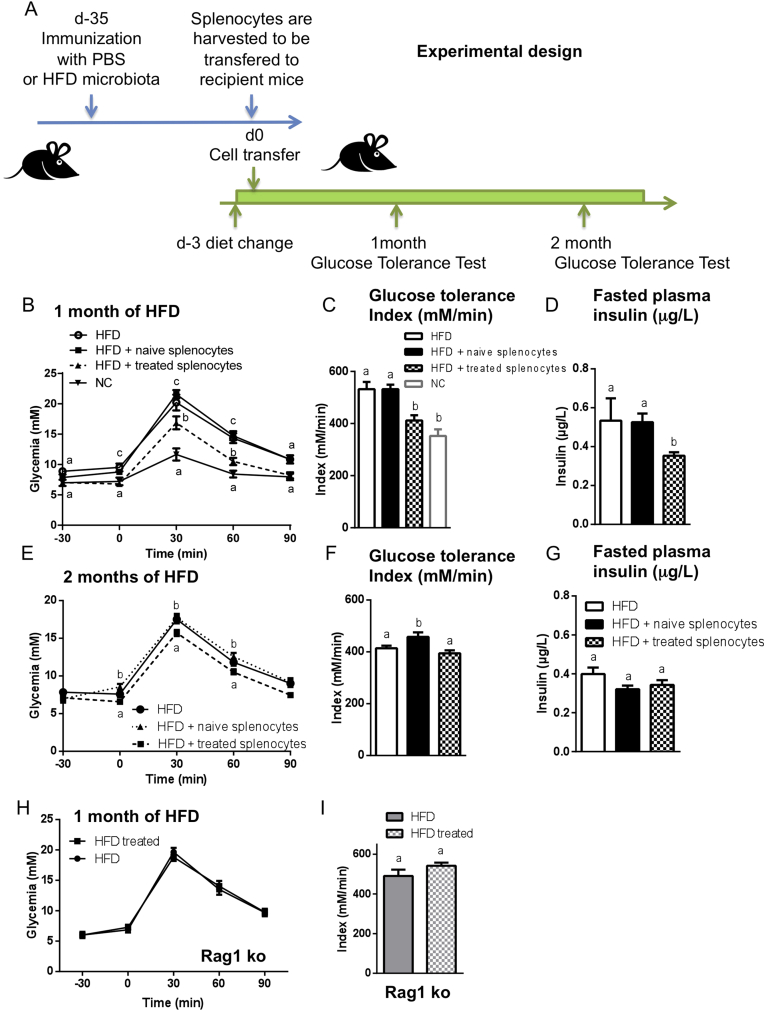
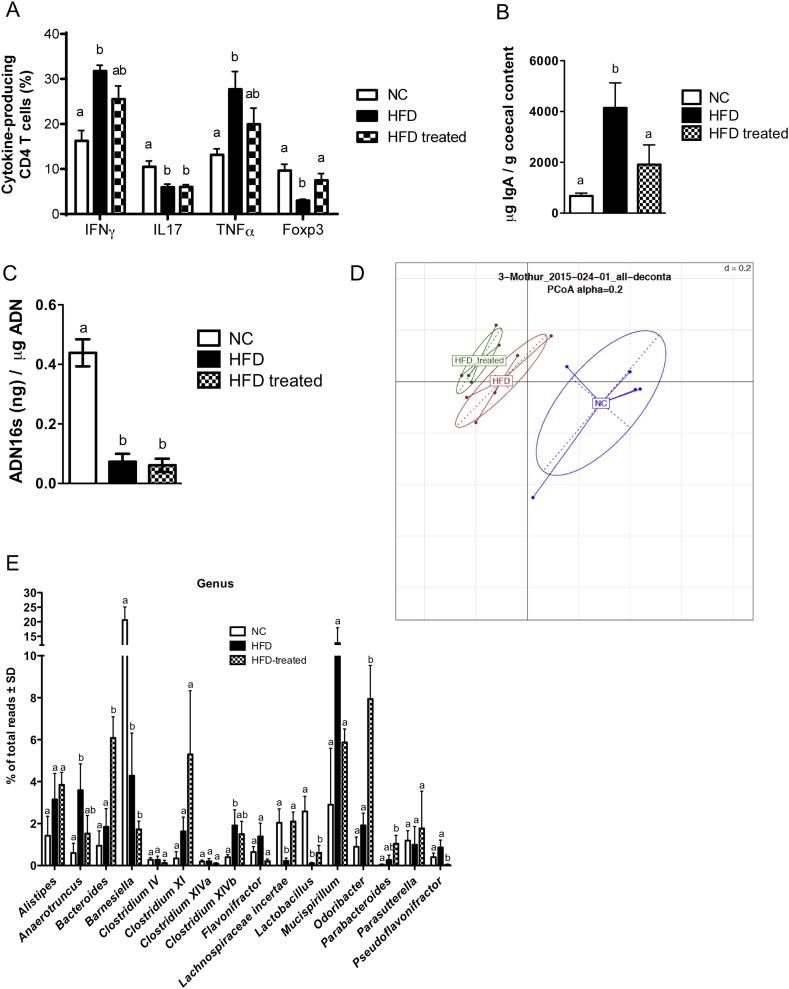
Methods: We triggered the immune system with microbial extracts specifically from the intestinal ileum contents of HFD-diabetic mice by the process of immunization. 35 days later, immunized mice were fed a HFD for up to two months in order to challenge the development of metabolic features. The immune responses were quantified. Eventually, adoptive transfer of immune cells from the microbiota-immunized mice to naïve mice was performed to demonstrate the causality of the microbiota-stimulated adaptive immune system on the development of metabolic disease. The gut microbiota of the immunized HFD-fed mice was characterized in order to demonstrate whether the manipulation of the microbiota to immune system interaction reverses the causal deleterious effect of gut microbiota dysbiosis on metabolic disease.
Results: Subcutaneous injection (immunization procedure) of ileum microbial extracts prevented hyperglycemia and insulin resistance in a dose-dependent manner in response to a HFD. The immunization enhanced the proliferation of CD4 and CD8 T cells in lymphoid organs, also increased cytokine production and antibody secretion. As a mechanism explaining the metabolic improvement, the immunization procedure reversed gut microbiota dysbiosis. Finally, adoptive transfer of immune cells from immunized mice improved metabolic features in response to HFD.
Conclusions: Glycemia and insulin sensitivity can be regulated by triggering the adaptive immunity to microbiota interaction. This reduces the gut microbiota dysbiosis induced by a fat-enriched diet.
Keywords: APC, Antigen presenting cells; AT, Adipose tissue; Gut microbiota and metabolic diseases; Immunity; Insulin resistance; LN, Lymph nodes; NC, Normal chow; T2D, Type 2 diabetes; VL, Vastus lateralis muscle.
Figures
Comment in
-
Diabetes: Gut microbiota improves dysglycaemia.Nat Rev Endocrinol. 2016 Jun;12(6):310. doi: 10.1038/nrendo.2016.57. Epub 2016 Apr 15. Nat Rev Endocrinol. 2016. PMID: 27080135 No abstract available.
References
-
- Qin J., Li Y., Cai Z., Li S., Zhu J., Zhang F. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature. 2012;490:55–60. - PubMed
-
- Ley R.E., Turnbaugh P.J., Klein S., Gordon J.I. Human gut microbes associated with obesity. Nature. 2006;444:1022–1023. - PubMed
-
- Turnbaugh P.J., Ley R.E., Mahowald M.A., Magrini V., Mardis E.R., Gordon J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444:1027–1031. - PubMed
-
- Garidou L., Pomie C., Klopp P., Waget A., Charpentier J., Aloulou M. The gut microbiota regulates intestinal CD4 T cells expressing RORgammat and controls metabolic disease. Cell Metabolism. 2015;22:100–112. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
