

Modulated Expression of Specific tRNAs Drives Gene Expression and Cancer Progression
- PMID: 27259150
- PMCID: PMC4915377
- DOI: 10.1016/j.cell.2016.05.046
Modulated Expression of Specific tRNAs Drives Gene Expression and Cancer Progression
Abstract
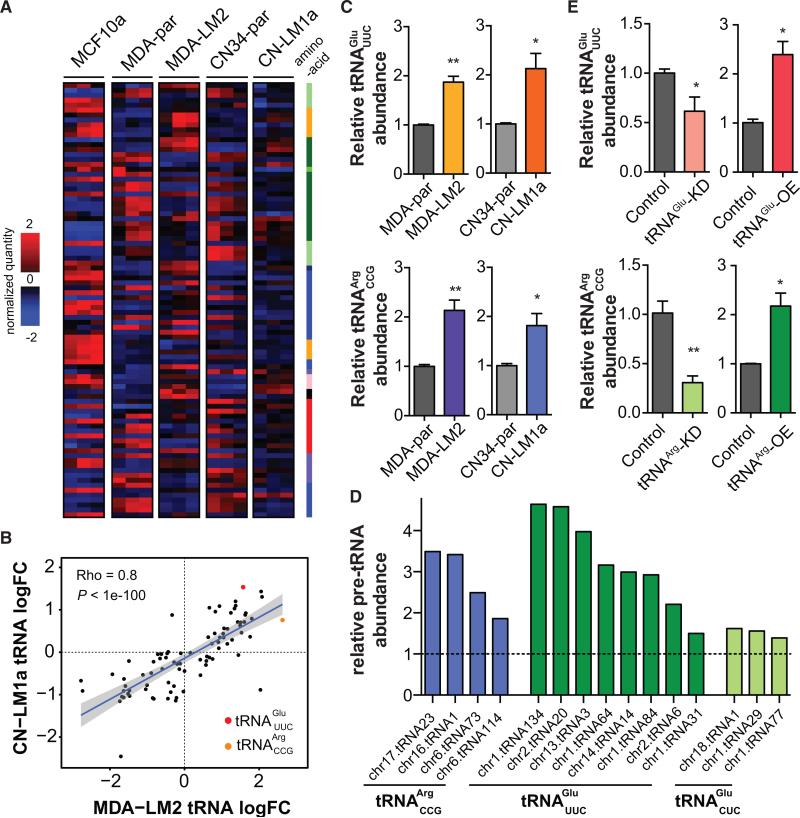
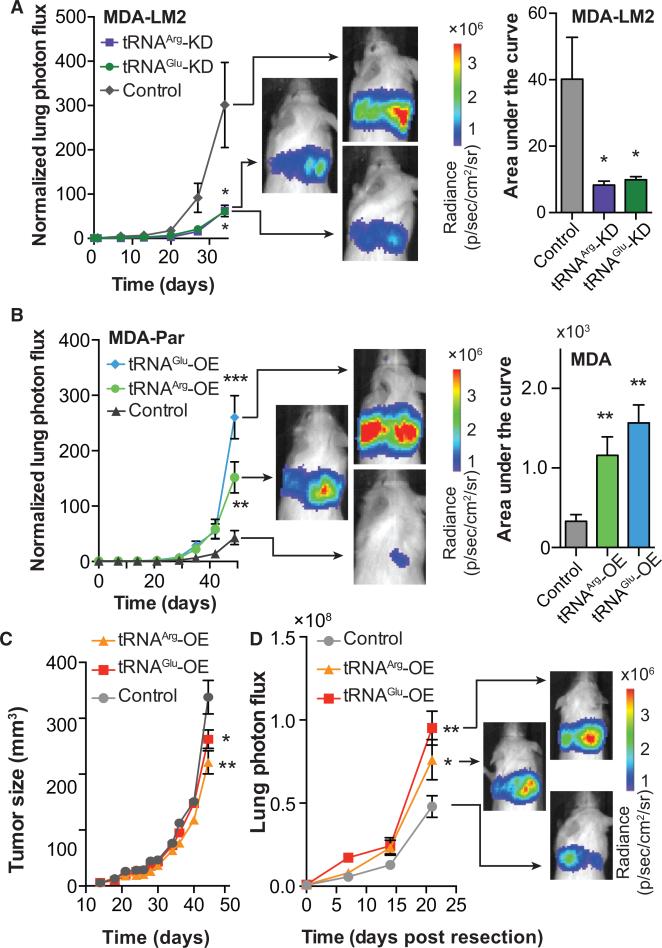
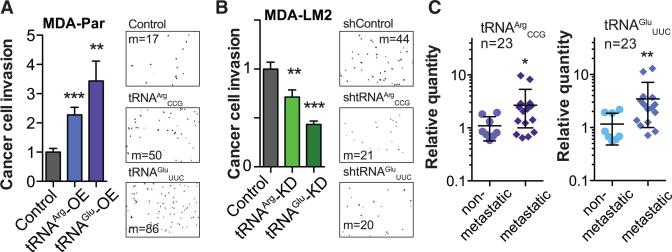
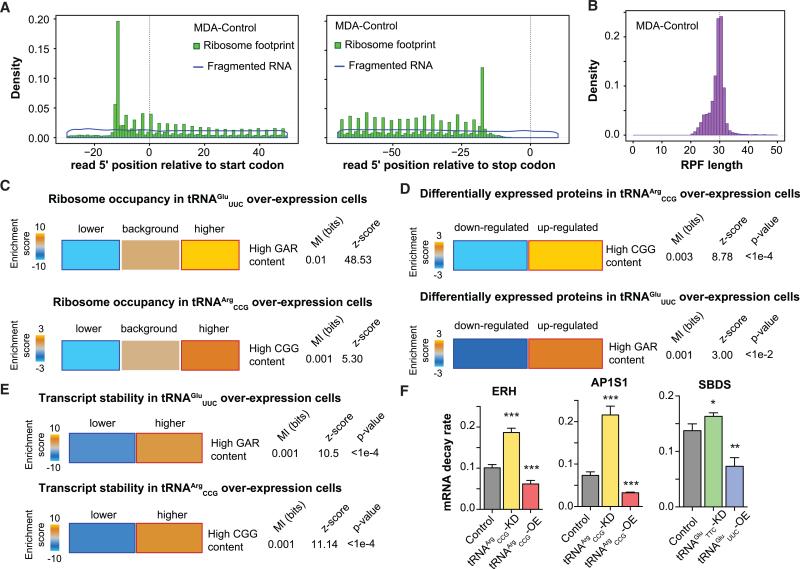
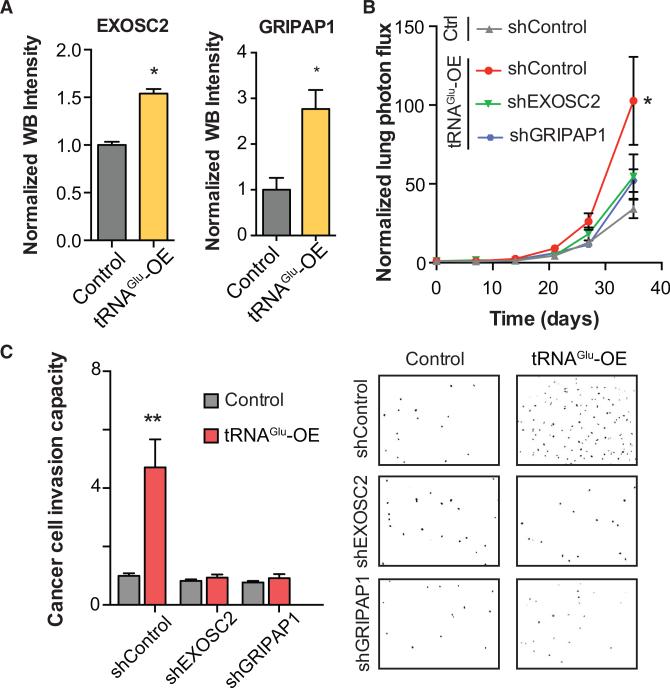
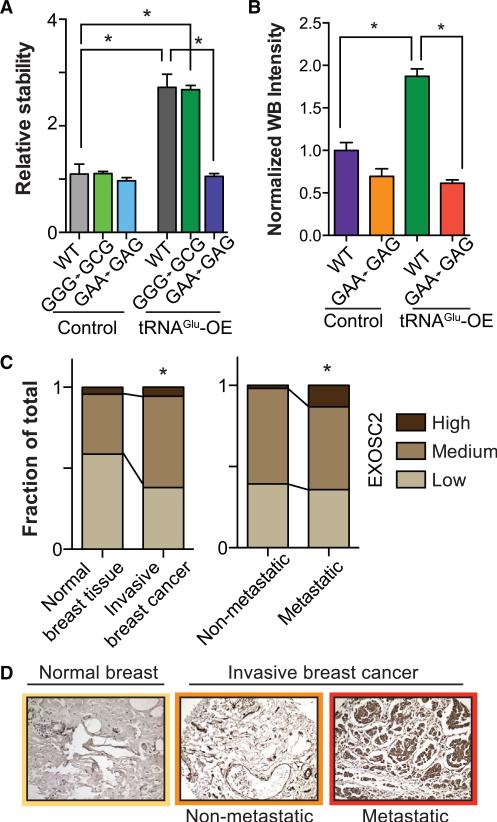
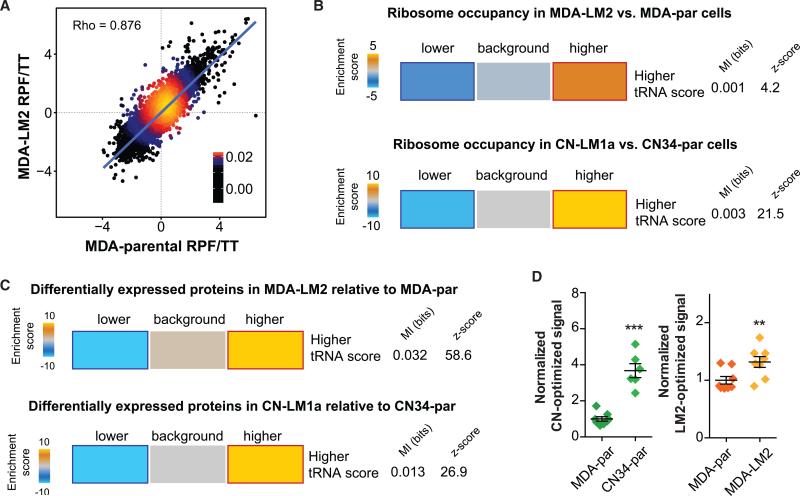
Transfer RNAs (tRNAs) are primarily viewed as static contributors to gene expression. By developing a high-throughput tRNA profiling method, we find that specific tRNAs are upregulated in human breast cancer cells as they gain metastatic activity. Through loss-of-function, gain-of-function, and clinical-association studies, we implicate tRNAGluUUC and tRNAArgCCG as promoters of breast cancer metastasis. Upregulation of these tRNAs enhances stability and ribosome occupancy of transcripts enriched for their cognate codons. Specifically, tRNAGluUUC promotes metastatic progression by directly enhancing EXOSC2 expression and enhancing GRIPAP1-constituting an "inducible" pathway driven by a tRNA. The cellular proteomic shift toward a pro-metastatic state mirrors global tRNA shifts, allowing for cell-state and cell-type transgene expression optimization through codon content quantification. TRNA modulation represents a mechanism by which cells achieve altered expression of specific transcripts and proteins. TRNAs are thus dynamic regulators of gene expression and the tRNA codon landscape can causally and specifically impact disease progression.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
A Pro-metastatic tRNA Pathway.Cell. 2016 Jun 2;165(6):1314-1315. doi: 10.1016/j.cell.2016.05.066. Cell. 2016. PMID: 27259143
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
