

ELAVL2-regulated transcriptional and splicing networks in human neurons link neurodevelopment and autism
- PMID: 27260404
- PMCID: PMC6086562
- DOI: 10.1093/hmg/ddw110
ELAVL2-regulated transcriptional and splicing networks in human neurons link neurodevelopment and autism
Abstract
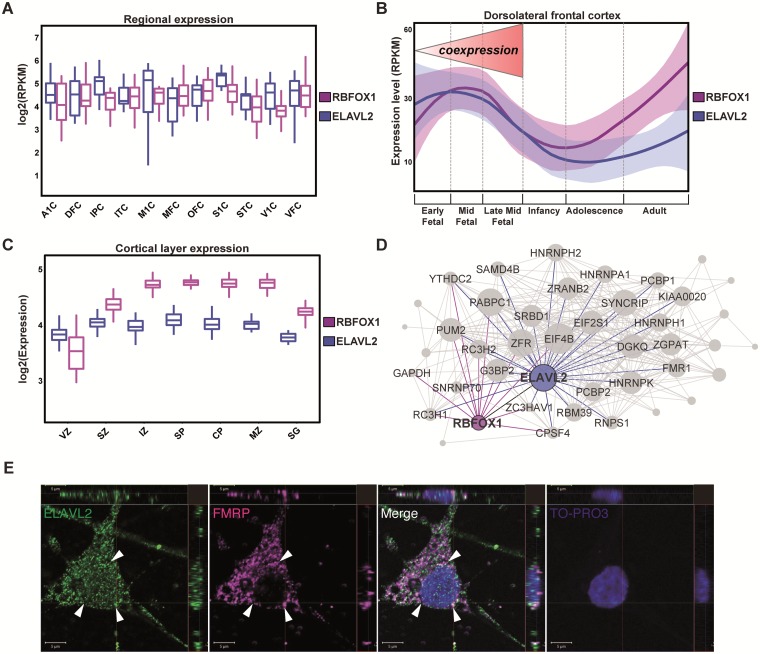
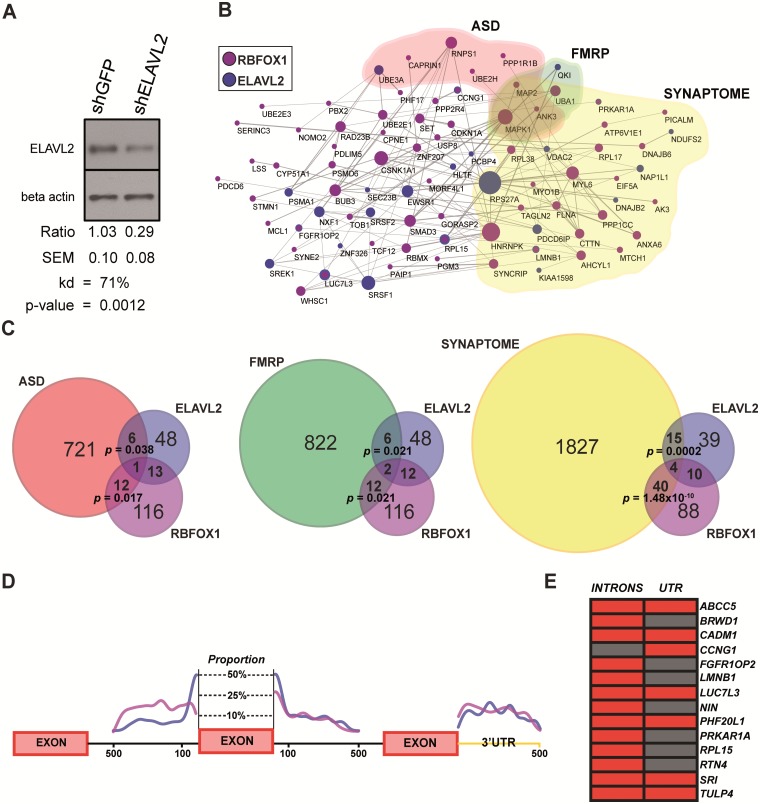
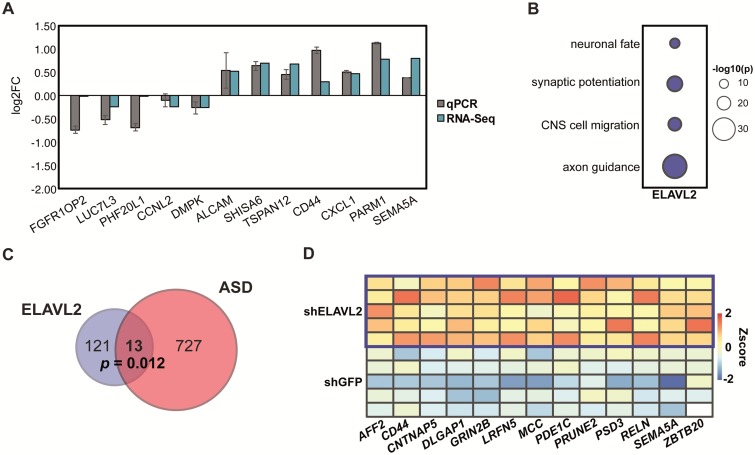
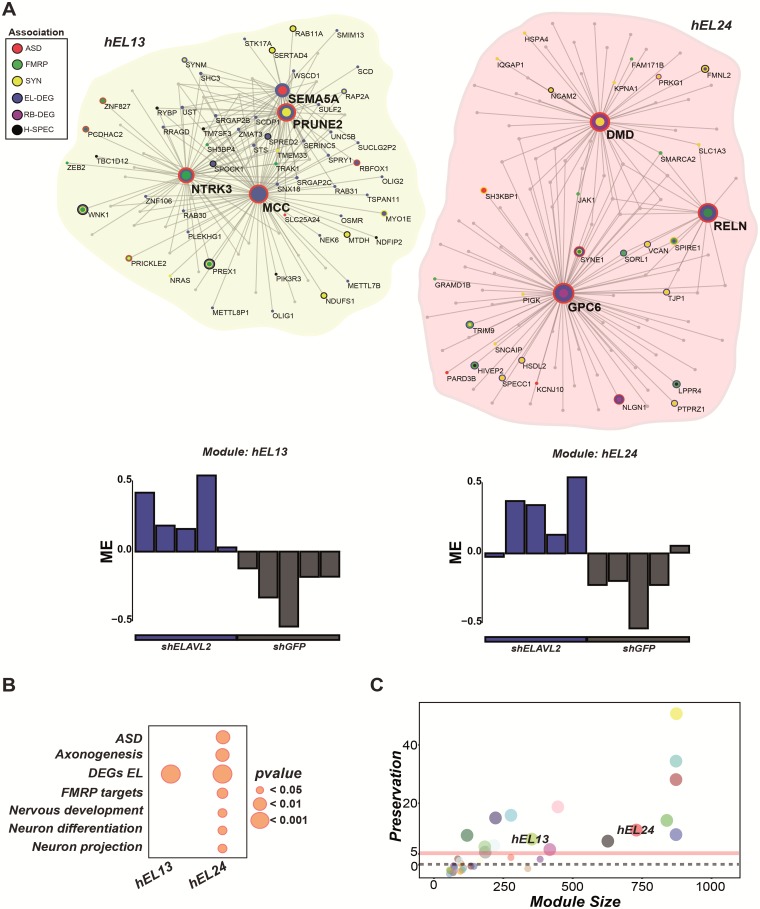
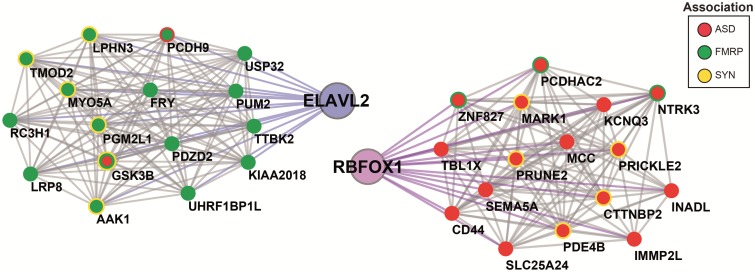
The role of post-transcriptional gene regulation in human brain development and neurodevelopmental disorders remains mostly uncharacterized. ELAV-like RNA-binding proteins (RNAbps) are a family of proteins that regulate several aspects of neuronal function including neuronal excitability and synaptic transmission, both critical to the normal function of the brain in cognition and behavior. Here, we identify the downstream neuronal transcriptional and splicing networks of ELAVL2, an RNAbp with previously unknown function in the brain. Expression of ELAVL2 was reduced in human neurons and RNA-sequencing was utilized to identify networks of differentially expressed and alternatively spliced genes resulting from haploinsufficient levels of ELAVL2. These networks contain a number of autism-relevant genes as well as previously identified targets of other important RNAbps implicated in autism spectrum disorder (ASD) including RBFOX1 and FMRP. ELAVL2-regulated co-expression networks are also enriched for neurodevelopmental and synaptic genes, and include genes with human-specific patterns of expression in the frontal pole. Together, these data suggest that ELAVL2 regulation of transcript expression is critical for neuronal function and clinically relevant to ASD.
© The Author 2016. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Raj B., Blencowe B.J. (2015) Alternative splicing in the mammalian nervous system: recent insights into mechanisms and functional roles. Neuron, 87, 14–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
