

Septate Junction Proteins Play Essential Roles in Morphogenesis Throughout Embryonic Development in Drosophila
- PMID: 27261004
- PMCID: PMC4978892
- DOI: 10.1534/g3.116.031427
Septate Junction Proteins Play Essential Roles in Morphogenesis Throughout Embryonic Development in Drosophila
Abstract
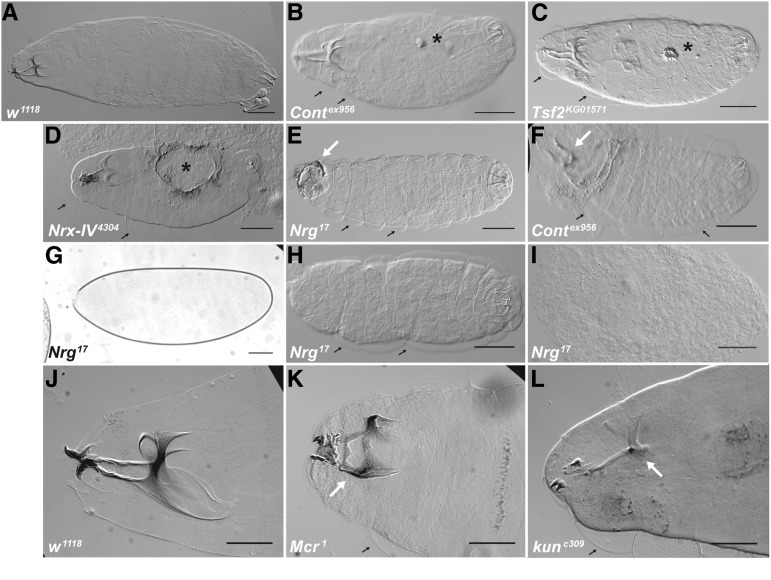
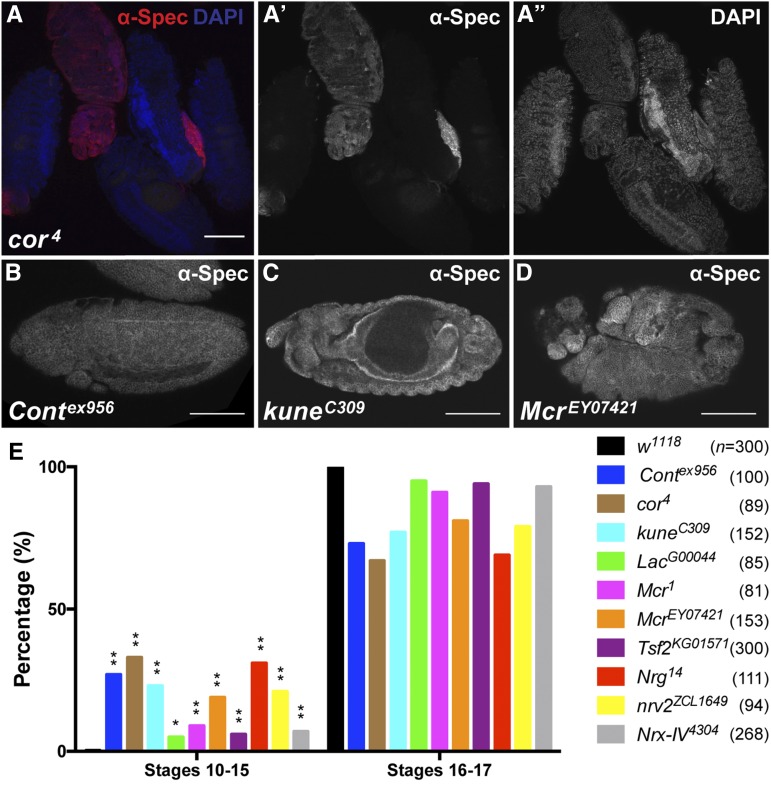
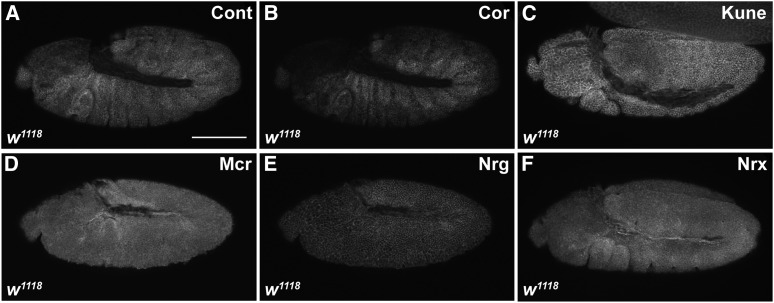
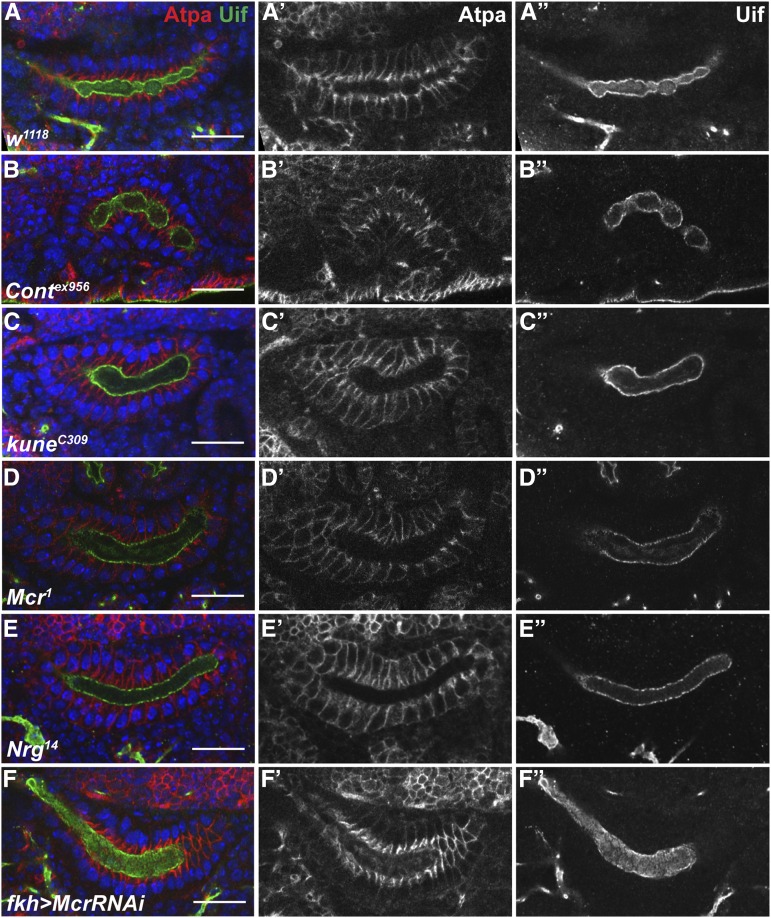
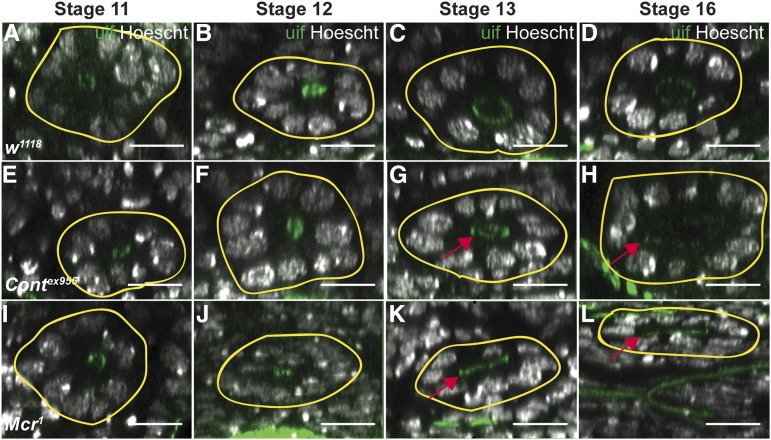
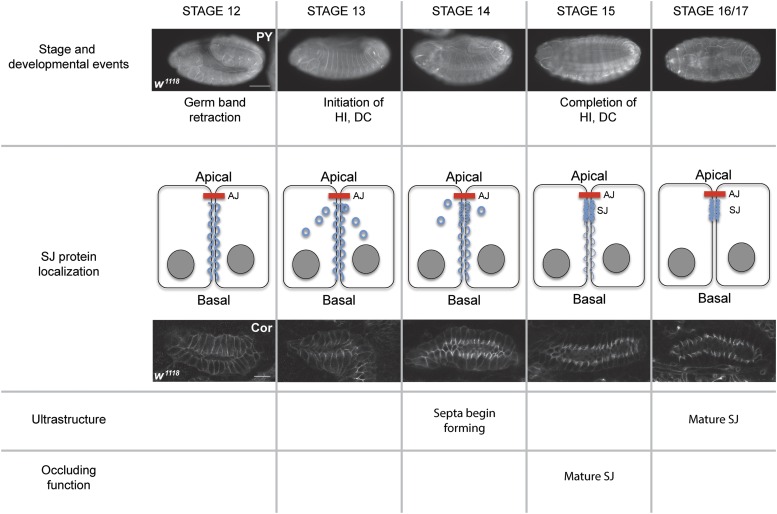
The septate junction (SJ) is the occluding junction found in the ectodermal epithelia of invertebrate organisms, and is essential to maintain chemically distinct compartments in epithelial organs, to provide the blood-brain barrier in the nervous system, and to provide an important line of defense against invading pathogens. More than 20 genes have been identified to function in the establishment or maintenance of SJs in Drosophila melanogaster Numerous studies have demonstrated the cell biological function of these proteins in establishing the occluding junction, whereas very few studies have examined further developmental roles for them. Here we examined embryos with mutations in nine different core SJ genes and found that all nine result in defects in embryonic development as early as germ band retraction, with the most penetrant defect observed in head involution. SJ genes are also required for cell shape changes and cell rearrangements that drive the elongation of the salivary gland during midembryogenesis. Interestingly, these developmental events occur at a time prior to the formation of the occluding junction, when SJ proteins localize along the lateral membrane and have not yet coalesced into the region of the SJ. Together, these observations reveal an underappreciated role for a large group of SJ genes in essential developmental events during embryogenesis, and suggest that the function of these proteins in facilitating cell shape changes and rearrangements is independent of their role in the occluding junction.
Keywords: dorsal closure; head involution; morphogenesis; salivary glands; septate junction.
Copyright © 2016 Hall and Ward.
Figures
Similar articles
-
Expanding the Junction: New Insights into Non-Occluding Roles for Septate Junction Proteins during Development.J Dev Biol. 2021 Mar 21;9(1):11. doi: 10.3390/jdb9010011. J Dev Biol. 2021. PMID: 33801162 Free PMC article. Review.
-
Septate junction proteins are required for egg elongation and border cell migration during oogenesis in Drosophila.G3 (Bethesda). 2021 Jul 14;11(7):jkab127. doi: 10.1093/g3journal/jkab127. G3 (Bethesda). 2021. PMID: 33871584 Free PMC article.
-
Septate junction proteins are required for cell shape changes, actomyosin reorganization and cell adhesion during dorsal closure in Drosophila.Front Cell Dev Biol. 2022 Sep 27;10:947444. doi: 10.3389/fcell.2022.947444. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36238688 Free PMC article.
-
Pasiflora proteins are novel core components of the septate junction.Development. 2015 Sep 1;142(17):3046-57. doi: 10.1242/dev.119412. Development. 2015. PMID: 26329602 Free PMC article.
-
Molecular organization and function of invertebrate occluding junctions.Semin Cell Dev Biol. 2014 Dec;36:186-93. doi: 10.1016/j.semcdb.2014.09.009. Epub 2014 Sep 17. Semin Cell Dev Biol. 2014. PMID: 25239398 Review.
Cited by
-
Expanding the Junction: New Insights into Non-Occluding Roles for Septate Junction Proteins during Development.J Dev Biol. 2021 Mar 21;9(1):11. doi: 10.3390/jdb9010011. J Dev Biol. 2021. PMID: 33801162 Free PMC article. Review.
-
Circulative Transmission of Cileviruses in Brevipalpus Mites May Involve the Paracellular Movement of Virions.Front Microbiol. 2022 Apr 6;13:836743. doi: 10.3389/fmicb.2022.836743. eCollection 2022. Front Microbiol. 2022. PMID: 35464977 Free PMC article.
-
Neuronal immunoglobulin superfamily cell adhesion molecules in epithelial morphogenesis: insights from Drosophila.Philos Trans R Soc Lond B Biol Sci. 2020 Oct 12;375(1809):20190553. doi: 10.1098/rstb.2019.0553. Epub 2020 Aug 24. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32829687 Free PMC article. Review.
-
Select Septate Junction Proteins Direct ROS-Mediated Paracrine Regulation of Drosophila Cardiac Function.Cell Rep. 2019 Aug 6;28(6):1455-1470.e4. doi: 10.1016/j.celrep.2019.07.004. Cell Rep. 2019. PMID: 31390561 Free PMC article.
-
Occluding junctions as novel regulators of tissue mechanics during wound repair.J Cell Biol. 2018 Dec 3;217(12):4267-4283. doi: 10.1083/jcb.201804048. Epub 2018 Sep 18. J Cell Biol. 2018. PMID: 30228162 Free PMC article.
References
-
- Batz T., Forster D., Luschnig S., 2014. The transmembrane protein Macroglobulin complement-related is essential for septate junction formation and epithelial barrier function in Drosophila. Development 141: 899–908. - PubMed
-
- Baumgartner S., Littleton J. T., Broadie K., Bhat M. A., Harbecke R., et al. , 1996. A Drosophila neurexin is required for septate junction and blood-nerve barrier formation and function. Cell 87: 1059–1068. - PubMed
-
- Behr M., Riedel D., Schuh R., 2003. The claudin-like megatrachea is essential in septate junctions for the epithelial barrier function in Drosophila. Dev. Cell 5: 611–620. - PubMed
-
- Bradley P. L., Myat M. M., Comeaux C. A., Andrew D. J., 2003. Posterior migration of the salivary gland requires an intact visceral mesoderm and integrin function. Dev. Biol. 257: 249–262. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases