

Visualizing retinoic acid morphogen gradients
- PMID: 27263412
- PMCID: PMC4933793
- DOI: 10.1016/bs.mcb.2016.03.003
Visualizing retinoic acid morphogen gradients
Abstract
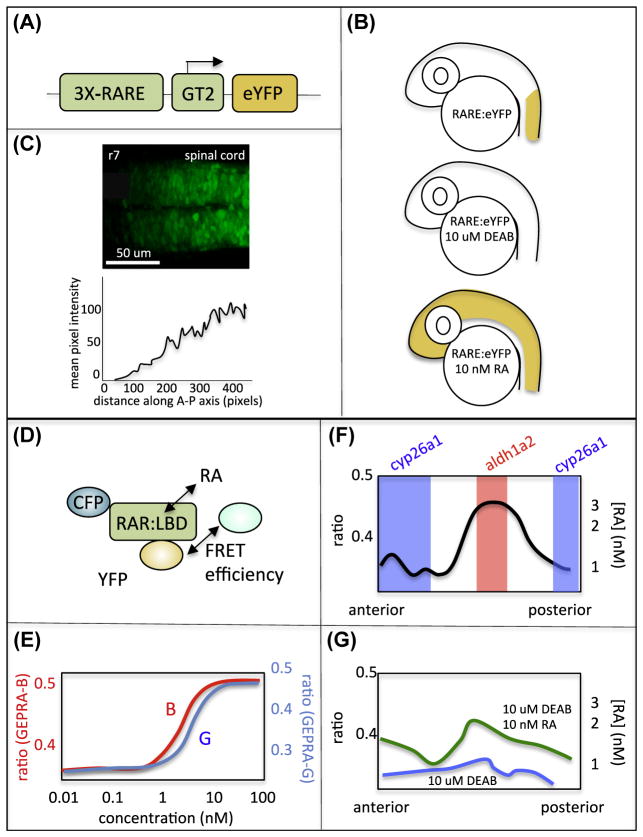
Morphogens were originally defined as secreted signaling molecules that diffuse from local sources to form concentration gradients, which specify multiple cell fates. More recently morphogen gradients have been shown to incorporate a range of mechanisms including short-range signal activation, transcriptional/translational feedback, and temporal windows of target gene induction. Many critical cell-cell signals implicated in both embryonic development and disease, such as Wnt, fibroblast growth factor (Fgf), hedgehog (Hh), transforming growth factor beta (TGFb), and retinoic acid (RA), are thought to act as morphogens, but key information on signal propagation and ligand distribution has been lacking for most. The zebrafish provides unique advantages for genetics and imaging to address gradients during early embryonic stages when morphogens help establish major body axes. This has been particularly informative for RA, where RA response elements (RAREs) driving fluorescent reporters as well as Fluorescence Resonance Energy Transfer (FRET) reporters of receptor binding have provided evidence for gradients, as well as regulatory mechanisms that attenuate noise and enhance gradient robustness in vivo. Here we summarize available tools in zebrafish and discuss their utility for studying dynamic regulation of RA morphogen gradients, through combined experimental and computational approaches.
Keywords: Diffusion; FRET; Morphogen; Retinoic acid; Rhombomere; Zebrafish.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures




Similar articles
-
Visualization of an endogenous retinoic acid gradient across embryonic development.Nature. 2013 Apr 18;496(7445):363-6. doi: 10.1038/nature12037. Epub 2013 Apr 7. Nature. 2013. PMID: 23563268
-
Noise modulation in retinoic acid signaling sharpens segmental boundaries of gene expression in the embryonic zebrafish hindbrain.Elife. 2016 Apr 12;5:e14034. doi: 10.7554/eLife.14034. Elife. 2016. PMID: 27067377 Free PMC article.
-
Cellular retinoic acid-binding proteins are essential for hindbrain patterning and signal robustness in zebrafish.Development. 2012 Jun;139(12):2150-5. doi: 10.1242/dev.077065. Development. 2012. PMID: 22619388 Free PMC article.
-
Dynamics and precision in retinoic acid morphogen gradients.Curr Opin Genet Dev. 2012 Dec;22(6):562-9. doi: 10.1016/j.gde.2012.11.012. Epub 2012 Dec 19. Curr Opin Genet Dev. 2012. PMID: 23266215 Free PMC article. Review.
-
Generation and interpretation of FGF morphogen gradients in vertebrates.Curr Opin Genet Dev. 2013 Aug;23(4):415-22. doi: 10.1016/j.gde.2013.03.002. Epub 2013 May 10. Curr Opin Genet Dev. 2013. PMID: 23669552 Review.
Cited by
-
Cell Identity Switching Regulated by Retinoic Acid Signaling Maintains Homogeneous Segments in the Hindbrain.Dev Cell. 2018 Jun 4;45(5):606-620.e3. doi: 10.1016/j.devcel.2018.04.003. Epub 2018 May 3. Dev Cell. 2018. PMID: 29731343 Free PMC article.
-
Regulating Retinoic Acid Availability during Development and Regeneration: The Role of the CYP26 Enzymes.J Dev Biol. 2020 Mar 5;8(1):6. doi: 10.3390/jdb8010006. J Dev Biol. 2020. PMID: 32151018 Free PMC article. Review.
-
Mechanisms of Feedback Regulation of Vitamin A Metabolism.Nutrients. 2022 Mar 21;14(6):1312. doi: 10.3390/nu14061312. Nutrients. 2022. PMID: 35334970 Free PMC article. Review.
-
Tracing anti-cancer and cancer-promoting actions of all-trans retinoic acid in breast cancer to a RARα epigenetic mechanism of mammary epithelial cell fate.Oncotarget. 2016 Dec 27;7(52):87064-87080. doi: 10.18632/oncotarget.13500. Oncotarget. 2016. PMID: 27894085 Free PMC article.
-
Mesoderm patterning by a dynamic gradient of retinoic acid signalling.Philos Trans R Soc Lond B Biol Sci. 2020 Oct 12;375(1809):20190556. doi: 10.1098/rstb.2019.0556. Epub 2020 Aug 24. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32829679 Free PMC article. Review.
References
-
- Alexander C, Zuniga E, Blitz IL, Wada N, Le Pabic P, Javidan Y, … Schilling TF. Combinatorial roles for BMPs and endothelin 1 in patterning the dorsal-ventral axis of the craniofacial skeleton. Development. 2011;138:5135–5146. http://dx.doi.org/10.1242/dev.067801. - DOI - PMC - PubMed
-
- Astrom A, Tavakkol A, Pettersson U, Cromie M, Elder JT, Voorhees JJ. Molecular cloning of two human cellular retinoic acid-binding proteins (CRABP). Retinoic acid-induced expression of CRABP-II but not CRABP-I in adult human skin in vivo and in skin fibroblasts in vitro. Journal of Biological Chemistry. 1991;266:17662–17666. - PubMed
-
- Balasubramanian R, Zhang X. Mechanisms of FGF gradient formation during embryogenesis. Seminars in Cell and Developmental Biology. 2015 http://dx.doi.org/10.1016/j.semcdb.2015.10.004. pii:S1084–9521(15)00189-5. - DOI - PMC - PubMed
-
- Begemann G, Schilling TF, Rauch GJ, Geisler R, Ingham PW. The zebrafish neckless mutation reveals a requirement for raldh2 in mesodermal signals that pattern the hindbrain. Development. 2001;128:3081–3094. - PubMed
