

Chalcone-based Selective Inhibitors of a C4 Plant Key Enzyme as Novel Potential Herbicides
- PMID: 27263468
- PMCID: PMC4893628
- DOI: 10.1038/srep27333
Chalcone-based Selective Inhibitors of a C4 Plant Key Enzyme as Novel Potential Herbicides
Abstract
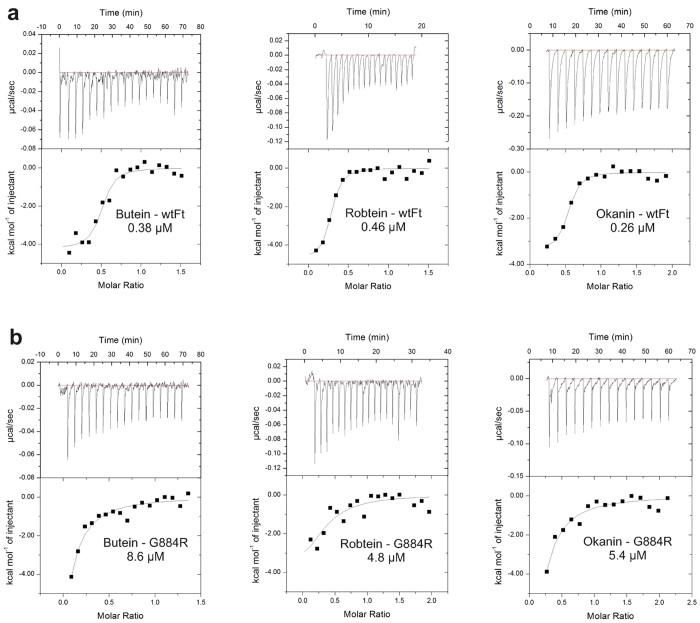
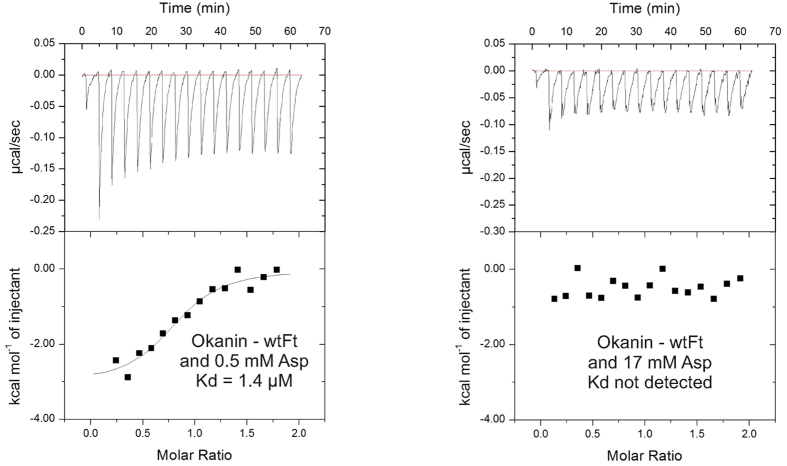
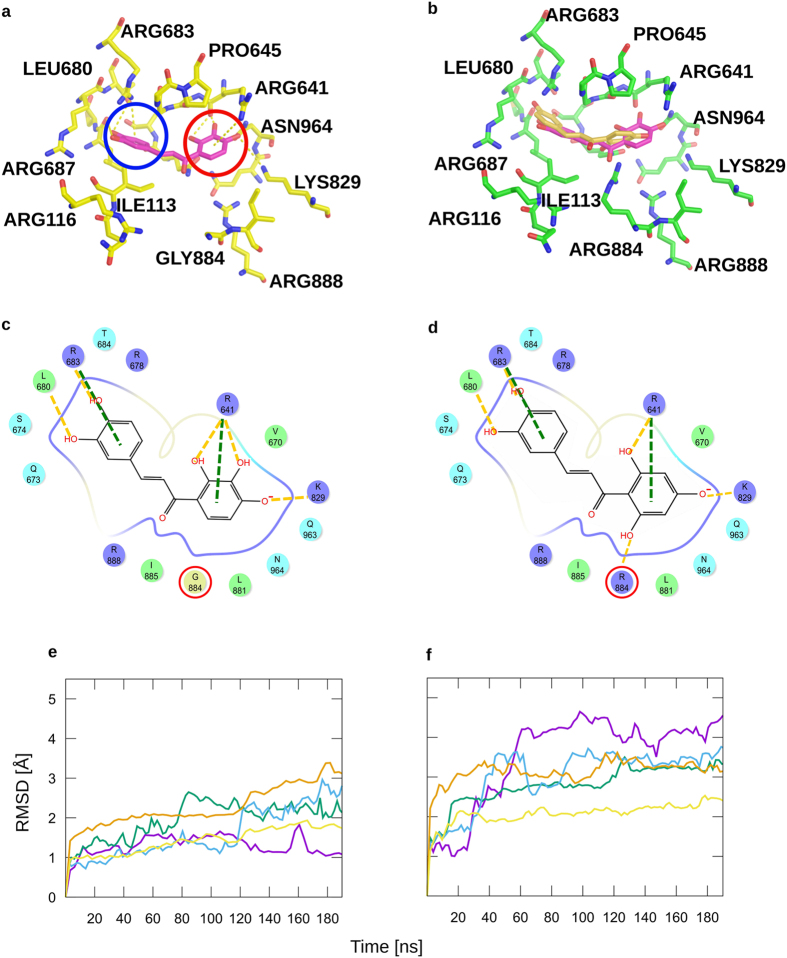
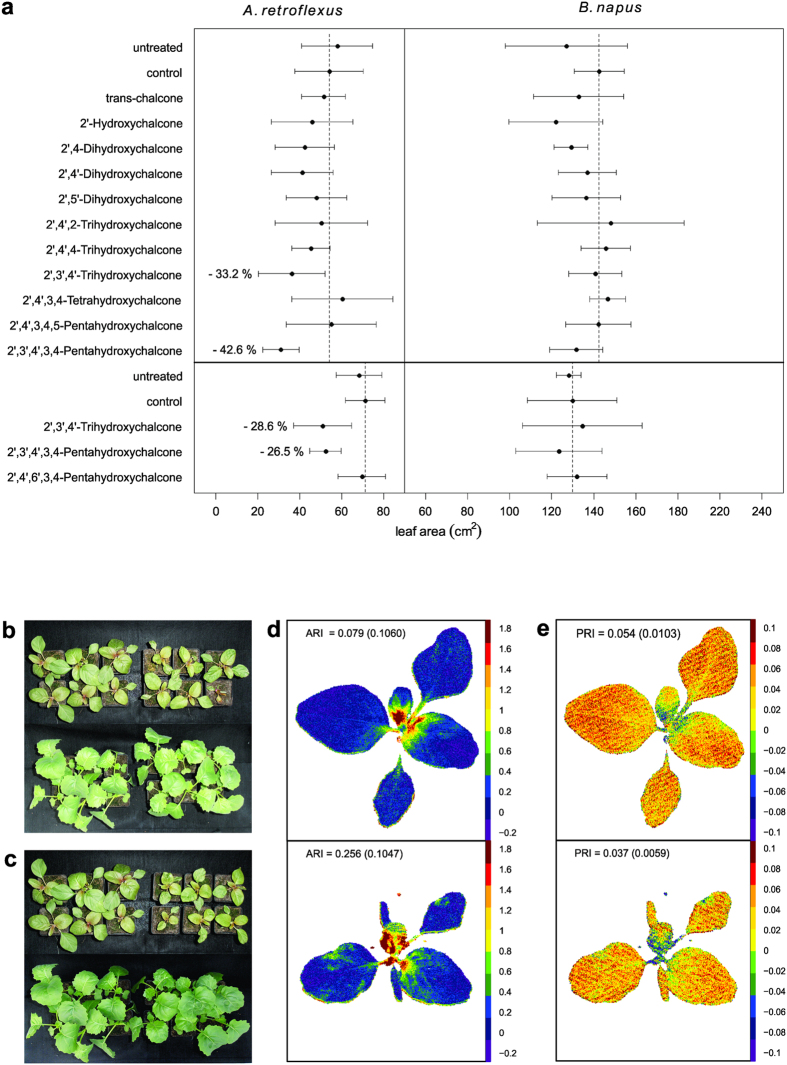
Weeds are a challenge for global food production due to their rapidly evolving resistance against herbicides. We have identified chalcones as selective inhibitors of phosphoenolpyruvate carboxylase (PEPC), a key enzyme for carbon fixation and biomass increase in the C4 photosynthetic pathway of many of the world's most damaging weeds. In contrast, many of the most important crop plants use C3 photosynthesis. Here, we show that 2',3',4',3,4-Pentahydroxychalcone (IC50 = 600 nM) and 2',3',4'-Trihydroxychalcone (IC50 = 4.2 μM) are potent inhibitors of C4 PEPC but do not affect C3 PEPC at a same concentration range (selectivity factor: 15-45). Binding and modeling studies indicate that the active compounds bind at the same site as malate/aspartate, the natural feedback inhibitors of the C4 pathway. At the whole plant level, both substances showed pronounced growth-inhibitory effects on the C4 weed Amaranthus retroflexus, while there were no measurable effects on oilseed rape, a C3 plant. Growth of selected soil bacteria was not affected by these substances. Our chalcone compounds are the most potent and selective C4 PEPC inhibitors known to date. They offer a novel approach to combat C4 weeds based on a hitherto unexplored mode of allosteric inhibition of a C4 plant key enzyme.
Conflict of interest statement
G.G. has patents pending on selective inhibition of C4 PEP carboxylases.
Figures
References
-
- Lepiniec L., Thomas M. & Vidal J. From enzyme activity to plant biotechnology: 30 years of research on phosphoenolpyruvate carboxylase. Plant Physiology and Biochemistry 41, 533–539 (2003).
-
- Sage R. F. The evolution of C4 photosynthesis. New Phytologist 161, 341–370 (2004). - PubMed
-
- Dong L.-Y., Masuda T., Kawamura T., Hata S. & Izui K. Cloning, Expression, and Characterization of a Root-Form Phosphoenolpyruvate Carboxylase from Zea mays: Comparison with the C4-Form Enzyme. Plant and Cell Physiology 39, 865–873 (1998). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
